小麦在我国已经有几千年的种植历史,种植区域几乎遍及全国各地。我国幅员辽阔,生态环境、气候环境、土壤类型和种植方式的不同,加之漫长的自然进化和人工选择,逐渐形成了丰富的小麦地方品种资源。在现代小麦遗传改良过程中,因为对产量的追求和少数骨干亲本品种资源的利用,导致小麦品种遗传基础日益狭窄,有些品种抗病虫害能力逐渐下降甚至丧失。小麦*粉白**病(Blumeria graminis f.sp. tritici)是危害小麦生产的主要病害之一,我国的小麦地方品种蕴含丰富的抗*粉白**病基因资源,一些地方品种具有良好的*粉白**病抗性。迄今,从我国小麦地方品种中发掘出的抗*粉白**病基因或等位基因包括Pm4e(D29)、Pm5d(PI 83395)、Pm5e(复壮30)、Pm24(齿牙糙)、P m24b(白葫芦)、Pm45(D57)、Pm47(红洋辣子)和Pm61(须须三月黄),另外还有多个临时命名的抗*粉白**病基因或等位基因,但是这些小麦地方品种来源的抗*粉白**病基因尚未被克隆。
为了解我国小麦地方品种对*粉白**病菌优势菌株的抗性,自2006年秋季我们开始对收集到的地方品种进行苗期抗病性鉴定,发现许多地方品种对*粉白**病菌株E09表现中抗或高抗,甚至免疫,其中来源于陕西的地方品种葫芦头表现高抗。之后王振忠同学在中国农业大学攻读博士学位期间(2010年9月–2015年6月)构建了石4185×葫芦头的遗传分离群体,利用E09菌株接种F1、F2代和F2:3家系进行遗传分析,发现葫芦头的*粉白**病抗性由显性单基因控制,暂时命名为MlHLT。通过BSA分析发现MlHLT与1DS染色体臂上的SSR标记Xgwm337连锁;进一步筛选找到了4个与MlHLT连锁的SSR标记Xgwm106、Xbarc119、Xcfd83和Xcfd72,将MlHLT定位于9.3 cM的遗传区间。为进一步加密MlHLT遗传区间,利用1DS染色体臂的EST序列开发多态性分子标记,获得与MlHLT紧密连锁的EST-STS标记Xwggc33,以此为基础寻找与该EST序列同源的粗山羊草SNP标记延伸序列、短柄草、水稻和高粱基因组序列中的同源基因,开展比较基因组分析,开发了多个多态性标记,将MlHLT锚定在分子标记Xwggc3026和Xwggc3148之间3.6 cM遗传区间,对应于粗山羊草基因组约13.4 Mb物理区间(Wang et al. 2015)。
由于来源于葫芦头(HLT)的抗*粉白**病基因MlHLT与地方品种齿牙糙(CYC)和白葫芦(BHL)中的抗*粉白**病基因Pm24(Huang et al. 1997)和Pm24b(Xue et al. 2012)均定位在小麦1DS染色体上大致相同的区间,于是我们进行了等位性测验,发现所有正反交组合[HLT×CYC(4,517 F2), HLT×BHL(4,318 F2), CYC×HLT(4,382 F2), CYC×BHL(4,590 F2)和BHL×HLT(4,692 F2)]的F2单株均高抗E09菌株,表明MlHLT、Pm24和Pm24b可能是等位基因或同一个基因(郭丽,2016)。随后利用93个全国各地采集的*粉白**病菌株对葫芦头、齿牙糙和白葫芦进行了抗性评价,发现这3个地方品种对全部菌株都表现抗病性(图1),且抗病反应型没有明显的差异,说明MlHLT、Pm24和Pm24b具有广谱*粉白**病抗性。因此克隆MlHLT、Pm24和Pm24b具有重要的理论和育种应用价值。

图1 葫芦头、齿牙糙、白葫芦和Chancellor对36个*粉白**病菌系的抗性表现
进一步利用粗山羊草的基因组序列信息开发多态性分子标记,使用3,720个单株的F2作图群体进行精细作图,将抗*粉白**病基因MlHLT定位在Xwggb 241和Xwgg b 244两个标记之间0.06 cM遗传区间,对应粗山羊草品系AL8/78的532 kb物理区间。序列分析和基因注释发现,MlHLT在中国春和粗山羊草的物理定位区间内均预测出6个候选基因:类受体激酶基因RLK1和RLK2、具有两个激酶结构域的小麦串联激酶基因(Wheat Tandem Kinase)WTK3、具有CC-NBS-LRR结构域基因CNL、假定蛋白HP和核糖体蛋白RP。分别从葫芦头和石4185中扩增这6个基因的全长DNA序列,发现仅有WTK3和CNL两个基因在葫芦头和石4185间存在序列差异,且在抗、感亲本苗期叶片中均表达,其序列差异均导致了所编码蛋白的氨基酸序列变异。与石4185相比,葫芦头中的WTK3基因在第3内含子处存在一个C/G的SNP变异,在第5外显子上存在一个6 bp的缺失,导致所编码蛋白有2个氨基酸(K400G401)的缺失;而石4185的CNL基因在第2外显子上有一处T/C的SNP引起的非同义氨基酸突变,在第3外显子上有一个单核苷酸缺失,分别引起编码蛋白的错义和无义突变(图2)。由于串联激酶基因WTK3和CNL基因都可能与抗病性相关,因此将二者均作为候选基因进行后续的功能验证。
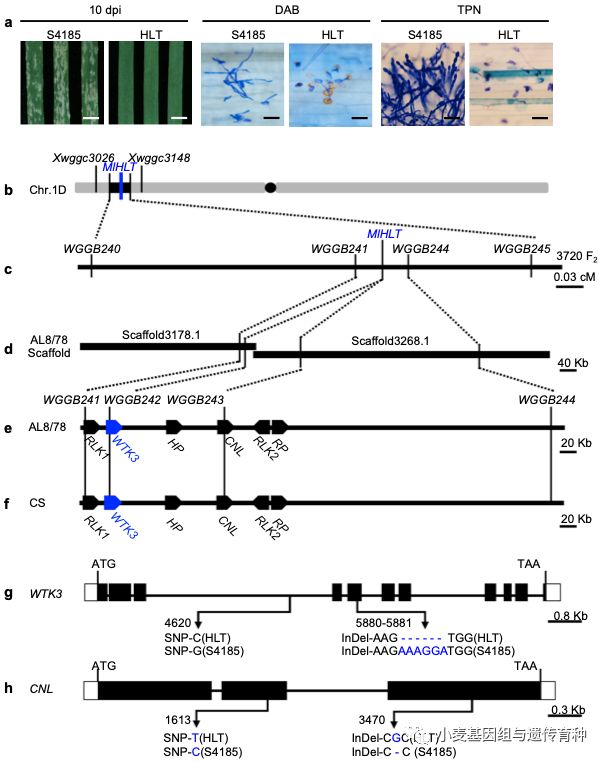
图2 小麦抗*粉白**病基因MlHLT图位克隆(Lu et al. 2020)
我们分别构建了WTK3基因自身启动子驱动基因组全长序列的互补载体和玉米Ubiquitin启动子驱动全长CDS的过表达载体,以及CNL基因自身启动子驱动基因组全长序列的互补载体(图3a),通过农杆菌介导的方法转化高感*粉白**病小麦品种Fielder,利用E09菌株接种T0代转基因植株和T1代家系,发现WTK3基因的互补载体和过表达载体转基因阳性植株均对*粉白**菌E09表现高抗,而CNL基因互补载体的所有转基因阳性植株都表现高感反应型(图3b),证明候选区间内的WTK3基因为MlHLT。
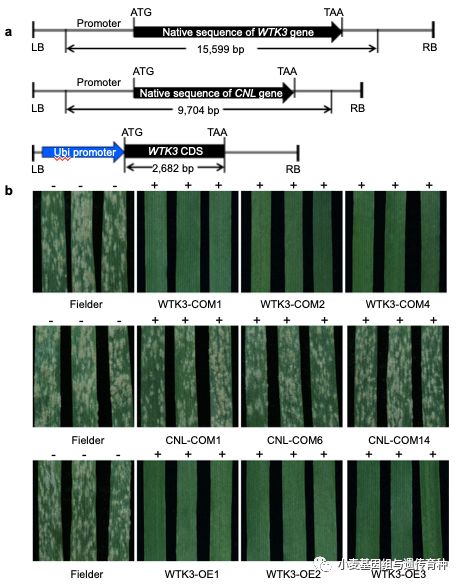
图3 小麦抗*粉白**病基因MlHLT转基因功能验证(Lu et al. 2020)
同时,利用EMS诱变葫芦头创制感病突变体,对M2代家系人工接种E09菌株进行筛选,获得了26个纯合感病突变体。对这26个感病突变体的WTK3和CN L基因全长DNA序列(包括启动子区和3’端序列)进行扩增测序,序列分析表明其中11个突变体在WTK3基因上发生了无义突变、错义突变(非同义氨基酸变异)和内含子-外显子拼接位点的改变(图4),在CNL基因上未发现突变位点。其中突变体Mut410和Mut1091的突变导致WTK3基因提前形成终止密码子;Mut1106的突变发生在WTK3基因第4内含子和第5外显子交界处,导致产生新的读码框发生移码突变;其余8个突变体的突变位点分别发生在WTK3基因的Kinase I和Kinase II两个激酶结构域上,且大部分突变发生在激酶的保守氨基酸残基上。突变体分析再次证明了WTK3为抗*粉白**病基因MlHLT。
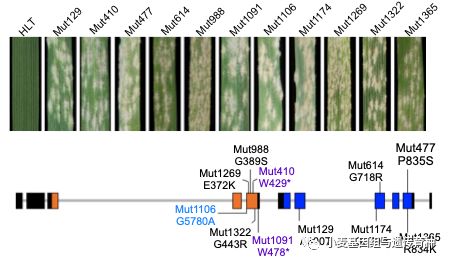
图4 小麦抗*粉白**病基因MlHLT突变体功能验证(Lu et al. 2020)
对齿牙糙和白葫芦中的WTK3基因序列进行了全长扩增,发现两个品种与葫芦头中的WTK3基因序列完全一致,说明MlHLT 、 Pm24和Pm24b为相同的基因。为了进一步了解Pm24基因在不同小麦种质资源中的序列变异,比较了7个粗山羊草和17个普通小麦中的WTK3基因全长基因组序列(10,410 bp),发现WTK3基因序列存在非常多的SNP和InDel变异。根据主要的序列变异特征可将这些种质的WTK 3基因划分为4个单倍型组(Group 1 – Group 4;图5),其中的第一个单倍型组(Group 1)包括葫芦头、齿牙糙、白葫芦、石4185和百农AK58等9个普通小麦品种(系),第二和第三单倍型组(Group 2和Group 3)均为粗山羊草,第四个单倍型组(Group 4)包括中国春等8个普通小麦品种(系)。与其它3个单倍型组相比,第一个单倍型组在WTK3基因的第7内含子上有一个1,509 bp的序列插入,然而WTK3基因第5外显子上的6 bp缺失(5880-AAAGGA-5881)只存在于第一个单倍型组的葫芦头、齿牙糙和白葫芦这3个地方品种中,其余21个小麦和粗山羊草品系均未发现这6 bp缺失。

图5 小麦抗*粉白**病基因WTK3全长序列单倍型组(Lu et al. 2020)
为了进一步明确WTK3基因6 bp缺失与其抗*粉白**病功能的关系,利用覆盖第5外显子该6 bp缺失区域632 bp序列的STS标记InDel-WTK3扩增了1,069份国内外多样化代表性种质,包括粗山羊草、中国小麦微核心种质、中国小麦地方品种、国外小麦地方品种等。序列比较发现该区域序列可分为10种单倍型(Hap I – Hap X; 图6),其中单倍型I(Hap I)仅包括葫芦头、齿牙糙、白葫芦和红芒麦4个中国小麦地方品种,这4份材料与其余1,065份材料的差异在于WTK3基因第5外显子上均存在6 bp缺失。序列分析发现红芒麦的WTK3基因全长基因组DNA和cDNA序列均与葫芦头、齿牙齿和白葫芦3个地方品种的WTK3基因完全一致,且对36个*粉白**病菌株都表现高抗反应型,表明红芒麦也含有Pm24基因。
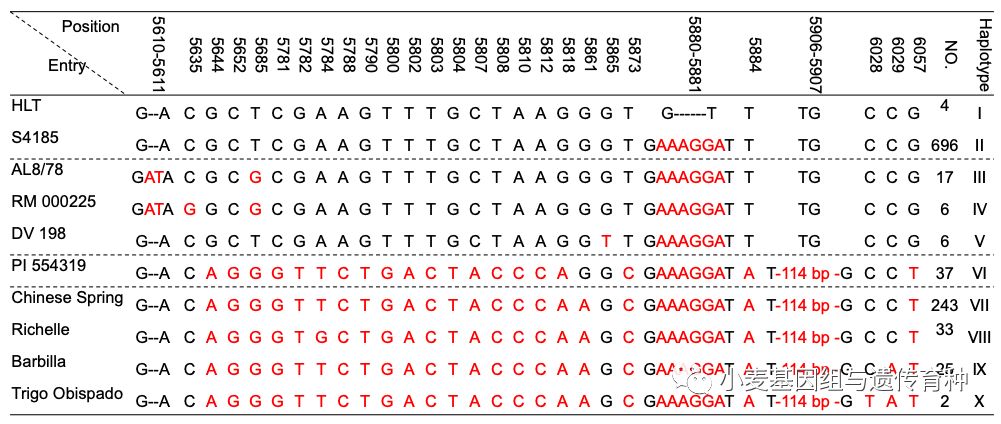
图6 小麦抗*粉白**病基因WTK3第5外显子区域序列单倍型(Lu et al. 2020)
WTK3 基因编码具有两个激酶结构域(Kinase I和Kinase II)的串联激酶蛋白(Tandem kinase protein, TKP),其中抗病等位基因WTK3HLT与感病等位基因WTK3S4185编码蛋白相比缺失了第400位的赖氨酸和401位的甘氨酸,这两个氨基酸的缺失发生在其预测结构的两个α螺旋之间的loop区域,而非在Kinase I结构域的保守氨基酸残基上。在自然界中,WTK3基因在其第5外显子上发生6 bp缺失后使WTK3HLT基因获得了抗*粉白**病功能(Gain of function),在发生缺失的位置未发现任何其它形式的变异。那么WTK3基因在其第5外显子上6 bp缺失部位缺失3 bp或缺失更多碱基,是否同样能赋予WTK3基因抗*粉白**病功能?为此,我们构建了另外3种过表达载体遗传转化Fielder,分别是ProUbi:WTK3#1(编码的蛋白缺失第400位的赖氨酸)、ProUbi:WTK3#2(编码的蛋白缺失第401位的甘氨酸)和ProUbi:WTK3#3(编码的蛋白缺失第399位的赖氨酸、第400位的赖氨酸、第401位的甘氨酸和第402位的色氨酸)(图7a)。结果发现ProUbi:WTK3#1、ProUbi:WTK3# 2和ProUbi:WTK3#3所有T1代转基因阳性植株都高感*粉白**病,只有缺失第400位赖氨酸和第401位甘氨酸两个氨基酸的ProUbi:WTK3HLT载体转基因阳性植株表现高抗,表明只有WTK3基因第5外显子上缺失这特定的两个氨基酸才能使其获得抗*粉白**病功能,缺失其中的一个或更多个氨基酸都不能赋予其抗*粉白**病功能。

图7 小麦抗*粉白**病基因WTK3第5外显子6 bp缺失与抗病功能相关(Lu et al. 2020)
串联激酶蛋白是一类新的参与植物免疫反应的激酶家族。目前在植物中发现的具有明确功能的串联激酶基因包括大麦抗秆锈病基因RPG1 (Brueggeman et al. 2002)、大麦抗散黑穗病基因Un8 (Zang et al. 2015)、小麦抗条锈病基因Yr15 (WTK1) (Klymiuk et al. 2018)、小麦抗秆锈病基因Sr60(WTK2) (Chen et al. 2019)和小麦抗*粉白**病基因Pm24(WTK3)。根据其两个激酶结构域的氨基酸残基序列保守性,WTK1蛋白是激酶-假激酶(Kinase-pseudokinase)结构、WTK2和Un8是激酶-激酶(Kinase-kinase)结构、RPG1是假激酶-激酶(Pseudokinase-kinase)结构。对WTK3蛋白的激酶结构域进行序列比较发现,WTK3蛋白的第一个激酶结构域(Kinase I)保守氨基酸残基与典型的激酶,如BIK1、Pto、Stpk-V、RPG1、WTK1和WTK2的激酶结构域保守序列相似,表明WTK3的第一个激酶可能具有激酶活性功能;而第二个激酶结构域(Kinase II)相应的氨基酸残基保守性较差,推测其与WTK1和RPG1中的假激酶结构域类似,可能为假激酶。有两个EMS感病突变体在第一个激酶功能域的关键保守氨基酸残基上发生了突变,说明WTK3蛋白的第一个激酶结构域对其*粉白**病抗性功能至关重要;但是也有部分EMS感病突变体在第二个激酶结构域的关键保守氨基酸出残基上或邻近氨基酸残基上发生了突变,表明第二个激酶结构域对WTK3蛋白的*粉白**病抗性功能也有重要作用。
进化分析发现,WTK1、WTK2和Un8可能来源于激酶结构域的融合,而RPG1和WTK3 可能来源于激酶结构域的重复(图8)。RPG1中Kinase I和Kinase II的氨基酸序列一致性为53.0%,而WTK3中Kinase I和Kinase II的氨基酸序列一致性仅为29.3%,表明WTK3中两个激酶结构域发生重复的时间相对较久远。WTK3在中国春和粗山羊草中都存在等位基因(TraesCS1D02G058900.1和AET1Gv20142700.38),在中国春和硬粒小麦Svevo的1B染色体上存在其部分同源基因(Homeologs或Homoeologs) (TraesCS1B02G075800.1和TRITD1Bv1G020600.1),但是在野生二粒小麦品系Zavitan的1A和1B染色体,中国春、硬粒小麦Svevo和乌拉尔图小麦G1812的1A染色体上均未发现WTK3的部分同源基因,很可能是在小麦的进化过程中A基因组上的WTK3部分同源基因发生了丢失。此外,在大麦和黑麦1H和1R染色体上存在WTK3的直系同源基因(Orthologs),在短柄草、水稻、玉米和高粱中均存在WTK3的同源基因(Homologs),但均位于非共线性的基因组区域。以上结果表明WTK3基因的产生可能发生在禾本科植物发生分化之前。
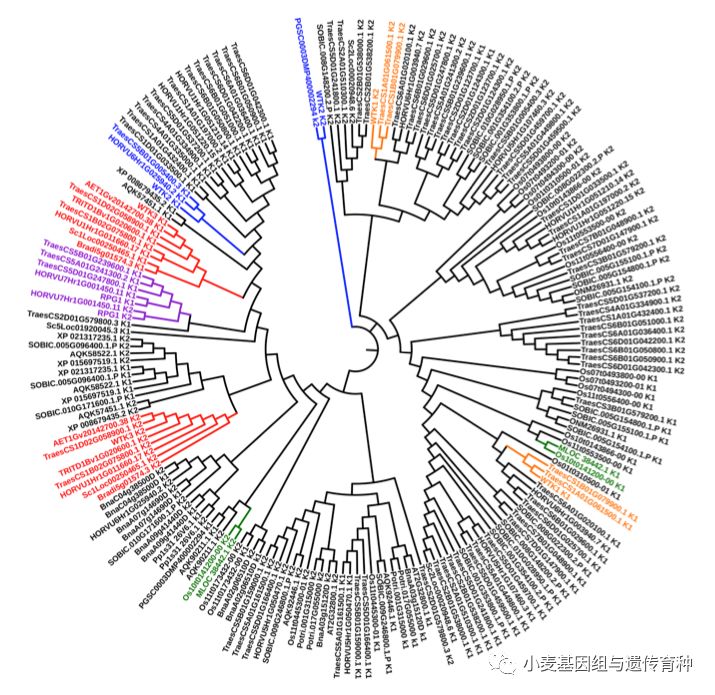
图8 小麦抗*粉白**病蛋白WTK3与植物蛋白激酶结构域的比较分析(Lu et al. 2020)
小麦抗*粉白**病基因Pm24编码的WTK3蛋白是首次发现具有两个激酶结构域的串联激酶具有抗*粉白**病功能。WTK3蛋白的两个氨基酸缺失赋予了Pm24基因的*粉白**病抗性,这种稀有的自然变异目前仅发现存在于中国小麦地方品种中。葫芦头、白葫芦、红芒麦和齿牙糙是收集自陕西和河南的小麦地方品种,从其地理分布推测是普通小麦引入中国之后在陕西关中平原及其邻近地区上千年的种植过程中,在病原菌选择压力下而产生的自然突变,之后被当地农民选择留存下来。抗病性鉴定和分子标记检测未在现代小麦育成品种中检测到含有Pm24基因,表明该优异基因在现代小麦育种中未能得到应用。该广谱抗*粉白**病基因的克隆使其在未来的小麦抗病性遗传改良,尤其是分子标记辅助选择和基因组编辑育种中具有较大的应用潜力。除Sr60外,目前已经发现的这些串联激酶基因均对其致病真菌具有广谱抗性,而且仅在麦类作物中发现串联激酶结构域基因具有抗病性功能,这可能预示着串联激酶所参与的植物抗病途径,或与病原菌的互作为麦类作物所特有。
小麦抗*粉白**病基因Pm24(WTK3)的克隆由中国科学院遗传与发育生物学研究所刘志勇课题组与中国农业科学院作物科学研究所李洪杰课题组合作完成,中国科学院遗传与发育生物学研究所刘翠敏课题组、中国农业大学、北京农学院、湖北省农业科学院植保土肥研究所俞大昭团队、美国UC Davis和USDA-ARS等单位的科学家参与了该项研究,研究结果于2020年2月3日在线发表于Nature Communications杂志(Lu et al. 2020)。
主要参考文献
-
Wang ZZ. et al. (2015) Genetic and physical mapping of powdery mildew resistance gene MlHLTin Chinese wheat landrace Hulutou.Theor. Appl. Genet. 128:365-373
-
Huang XQ. et al. (1997) Chromosomal location of genes for resistance to powdery mildew in common wheat (Triticum aestivumL. em. Thell.) 4. GenePm24in Chinese landrace Chiyacao.Theor. Appl. Genet. 95:950-953
-
Xue F. et al. (2012) Molecular mapping of a powdery mildew resistance gene in common wheat landrace Baihulu and its allelism with Pm24.Theor. Appl. Genet. 125:1425-1432.
-
郭丽 (2016) 小麦抗*粉白**病基因MlHLT精细定位和候选基因克隆. 中国农业大学硕士学位论文
-
Brueggeman R. et al. (2002) The barley stem rust-resistance gene Rpg1is a novel disease-resistance gene with homology to receptor kinases.Proc. Natl. Acad. Sci. USA 99:328-9333
-
Zang W. et al. (2015) Fine mapping and identification of a candidate gene for the barley Un8true loose smut resistance gene.Theor. Appl. Genet. 128:1343-1357
-
Klymiuk V. et al. (2018) Cloning of the wheat Yr15resistance gene sheds light on the plant tandem kinase-pseudokinase family.Nat. Commun. 9:3735
-
Chen S. et al. (2019) Wheat gene Sr60encodes a protein with two putative kinase domains that confers resistance to stem rust.New Phytol. DOI:10.1111/nph.16169
-
Lu P. et al. (2020) A rare gain of function mutation in a wheat tandem kinase confers resistance to powdery mildew. Nat . Commun. DOI: 10.1038/s41467-020-14294-0
