
在阅读此文之前,请点击一下“关注”,既方便您进行讨论和分享,又能给您带来不一样的参与感,感谢您的支持
文|Read
编辑|Read
前言
Polycomb group 蛋白是表观遗传转录抑制因子,调控许多发育过程,并被认为参与胚胎干细胞状态的维持。

利用CRISPR-Cas9技术产生具有单一或联合缺失Pcgf3和Pcgf5的胚胎干细胞。这些突变细胞仍保持自我更新和形成集落的能力,在体外和体内均显示出严重的中胚层分化缺陷。
RNA-seq分析单一或联合Pcgf3/5缺陷ES细胞的转录谱,发现, 与相关的多能性转录抑制复合物1在基因抑制中的传统作用相反 ,Pcgf3/5主要作为转录激活因子,在驱动许多参与中胚层分化的基因表达方面发挥作用。

蛋白组学方法和启动子占有率分析有助于建立一个扩展的Pcgf3/5相互作用网络,鉴定了几个新的Pcgf3/5相互作用蛋白。其中蛋白10,它可能转录共激活因子p300直接促进转录激活。
ES细胞中的Pcgf3/5缺失显著降低了Tex10和p300在目标基因上的结合。证明Pcgf3/5对ES细胞中组蛋白修饰因子H2AK119ub1的全局水平调控是必要的。Pcgf3/5在ES细胞中作为转录激活因子与Tex10和p300相互作用,并指出Pcgf3/5在多能性维持中具有冗余活性。

胚胎干细胞是一类具有自我更新和分化为体内几乎所有细胞系的能力的细胞
PcG蛋白是参与ES细胞多能性和分化的重要表观遗传调节因子。PcG最初被描述为果蝇中控制正确体节分割的一组基因, 维持Hox基因表达的转录抑制状态实现其功能。
在哺乳动物中,PcG蛋白形成至少两个不同的蛋白质复合物,即PRC1和PRC2,介导发育基因的沉默。
PRC2由核心亚单位Eed、Suz12和Ezh1/2组成,并对组蛋白H3的赖氨酸27位进行二甲基或三甲基化。PRC1则包含E3连接酶Ring1A/B, 用于对组蛋白H2A的赖氨酸119位进行单一泛素化。

PRC1可以根据其亚单位组成分为经典和非经典PRC1,经典PRC1含有Cbx蛋白Phc1/2/3、Pcgf2/4和Ring1A/B,而非经典PRC1则含有Rybp或Yaf2代替Cbx蛋白。
PRC1还根据Ring1A/B与Pcgf1–6的互斥结合进一步多样化,共有至少六个不同的PRC1复合物,即PRC1.1–1.6,每个复合物包含其中一个Pcgf。
Pcgf3和Pcgf5被确定为非经典PRC1.3和PRC1.5的关键组分,在293T-REx细胞*共中**享Ring1A/B、Rybp/Yaf2、激酶Ck2和Auts2,一些PcG复合物作为转录激活因子发挥作用, 表明最初对PcG蛋白的分类为转录抑制因子可能不适用于所有情况。

PRC1.5的组分Auts2能招募Ck2并与p300相互作用,使该复合物具有激活转录的能力。与哺乳动物Pcgf3同源的果蝇蛋白L73Ah参与调控果蝇中全局H2AK118ub1水平, 形成与Ring1的二聚体,Pcgf3和Pcgf5在哺乳动物中的功能尚待进一步确定。
CRISPR/Cas9基因工程技术产生了具有单一或联合Pcgf3/5缺失的ES细胞。Pcgf3或Pcgf5突变细胞是可行的并保留其典型的未分化ES细胞形态,Pcgf3或Pcgf5的缺失会影响ES细胞适当地分化为中胚层的能力, 无论是在体外还是在体内。

采用生化亲和纯化技术纯化了ES细胞中的Pcgf3/5蛋白复合物,并质谱分析进行了进一步分析。发现Pcgf3/5能够形成新型的非经典PRC1复合物,其中包含激酶Ck2和Tex10。展示了与Tex10相互作用,p300被招募到Pcgf3/5的靶基因上,可能有助于转录激活。
Pcgf3/5对ES细胞的自我更新并非必需
当Pcgf3和Pcgf5在维持ES细胞中的功能时,利用CRISPR/Cas9技术生成了Pcgf3或Pcgf5敲除的ES细胞系。为了删除编码RING指环结构域的区域并引入移码,设计了两个特异性sgRNA, 分别夹挟小鼠Pcgf3或Pcgf5基因的外显子2。

Western印迹分析显示目标基因敲除突变ES细胞中完全丧失了Pcgf3或Pcgf5蛋白,未检测到截短蛋白。Pcgf3和Pcgf5在氨基酸上有57.9%的同源性,并且在具有类似亚单位组成的复合物中存在。
单一或双重敲除Pcgf3/5的ES细胞是可行的,并且在被播种于丝裂霉素-C辐射的小鼠胚胎成纤维细胞上后,它们仍保留了正常形成典型未分化ES细胞集落的能力。 不同突变体的集落大小与野生型相当。
对多能性因子进行的免疫印迹分析表明,在ES细胞中敲除Pcgf3、Pcgf5或两者并没有显著改变核心多能性标志物Oct4、Nanog和Sox2的蛋白水平,与观察到的这些突变体的未分化形态一致。
单独或同时缺乏Pcgf3/5的ES细胞的四个独立衍生克隆获得了相同的表型。缺乏PRC1.2和PRC1.4的经典PRC1复合物核心成分Pcgf2或Pcgf4的ES细胞观察到非常相似的表型。除了Pcgf6, 在ES细胞的自我更新中是可有可无的。

Pcgf1的缺失严重影响了ES细胞的体外分化,主要是由于未能启动适当的分化程序, 而Pcgf6突变体ES细胞在EB中向内胚层系列分化出现偏倚。

将突变体和野生型ES细胞在没有滋养层细胞抑制因子的条件下培养,使其在悬滴中聚集形成EB。野生型、Pcgf3/5单一和双重基因敲除ES细胞在悬滴中培养3天后,继续在没有LIF的旋转条件下培养, 使细胞分化为三个胚层。
突变ES细胞保留了形成与对照细胞相似大小的EB的能力,在培养3、7和12天后,从ES细胞和EB中提取总RNA,并进行定量实时RT-PCR分析核心多能性基因转录水平以及特定于胚层的分化标记物。

核心多能性标志物Oct4和Nanog的表达被有效下调,而分化特异性标志物,包括内胚层(Gata6和Foxa2)、外胚层(Fgf5和Nestin)和中胚层(Brachyury,Flk1和Nkx2-5)的标志物,在分化过程中上调,与突变体和野生型细胞中分化程序的开始相吻合。值得注意的是,整个突变体EB培养过程中,与对照EB相比,所有突变体的中胚层标志物表达水平显著降低,而在内胚层和外胚层标志物的水平上未观察到突变体和野生型之间的差异, Pcgf3/5在EB培养过程中对于适当的中胚层系列分化是必要的。

Pcgf3/5双重基因敲除EB显示出比任何相应单突变体更明显的中胚层标志物表达降低,表明Pcgf3/5在EB分化过程中具有部分冗余功能。还检查了分化EB中这些胚层标志物的蛋白水平,发现在分化EB的第12天,中胚层标志物的蛋白水平与对照相比显著下降。
在内胚层和外胚层标志物的蛋白水平上未观察到突变体和野生型之间的差异,携带FLAG标记的Pcgf3/5的表达能够将中胚层标志物的蛋白水平恢复到与对照细胞相似的水平, 表明观察到的分化表型是由于Pcgf3/5缺陷引起的。
使用特异性抗α-平滑肌肌动蛋白的免疫组织化学,显示野生型ES细胞来源有大量SMA阳性细胞, 而来源于Pcgf3/5单一或联合缺失的ES细胞的SMA阳性细胞显著减少。
Pcgf3/5的消失对于内胚层和外胚层的规格没有影响,Pcgf3和Pcgf5在ES细胞体外和体内的中胚层分化中具有冗余必要的作用。

Pcgf3/5在ES细胞中积极调节转录活动
在Pcgf3缺失和野生型ES细胞之间比较,确定了318个差异表达基因,根据2倍差异标准进行筛选,而在Pcgf5缺失和野生型ES细胞之间有279个差异表达基因,Pcgf3−/−和Pcgf5−/−的基因列表也在很大程度上重叠。
对Pcgf3/5双突变体的转录谱分析显示,在双突变体中,显著差异表达的基因数量比Pcgf3/5单突变体增加了30%。 与野生型相比,双突变体中显著差异表达的基因中,>70%为下调基因。

根据基因本体术语分析,Pcgf3/5抑制的基因没有特定功能类别或细胞过程的显著富集。对Pcgf3−/−或Pcgf5−/−的下调基因,尤其是Pcgf3/5双突变体,进行GO分析显示它们与中胚层分化相关基因,暗示了Pcgf3/5在中胚层细胞命运中的特殊作用。
这些结果强烈表明Pcgf3和Pcgf5主要作为ES细胞的转录激活因子。不仅支持Pcgf3和Pcgf5在调控类似下游基因上具有相似作用的结论, 还为Pcgf3和Pcgf5在ES细胞分化中的冗余提供了分子解释。

之前的蛋白质组学研究确定Pcgf3和Pcgf5是非典型PRC1复合物的组成部分, 分别被称为PRC1.3或PRC1.5, 在293T-REx细胞中包含与Ring1B相关的Ck2、Rybp和Auts2。
ES细胞中PRC1.3或PRC1.5复合物的组成,在Pcgf3−/−或Pcgf5−/− ES细胞中稳定表达带有C端FLAG标签的Pcgf3或Pcgf5,并从核提取物中纯化它,然后利用质谱分析确定相关的蛋白质。

Pcgf5与Auts2和293T-Rex细胞中相互作用,在Pcgf3/5免疫沉淀物中检测到任何Auts2。Pcgf3和Pcgf5是互斥的,因为Pcgf3的免疫沉淀未能回收Pcgf5,反之亦然。
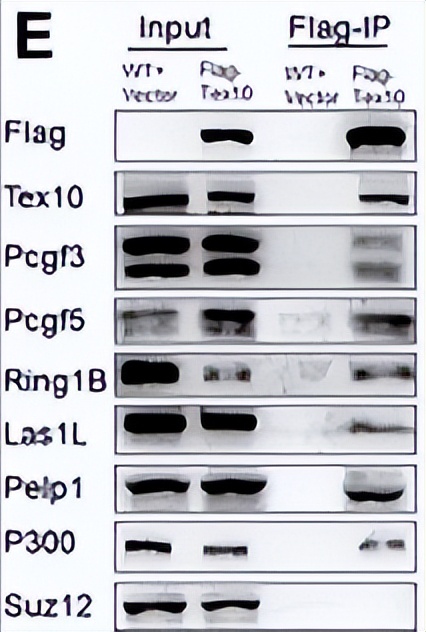
在重表达带有FLAG标签的Pcgf3或Pcgf5的突变ES细胞的核提取物中进行免疫沉淀实验,并使用FLAG抗体确认Pcgf3或Pcgf5与Ring1A/B、Rybp、Ck2s、Tex10、Las1L和Pelp1的结合。

未与PRC1.3或PRC1.5相关的其他PcG组分,分别组成PRC1.6和PRC2, 与Pcgf3或Pcgf5不沉淀。
在稳定表达带有FLAG标签的Ck2s的野生型ES细胞中,Pcgf3/5但不是Pcgf6蛋白质存在于被抗-FLAG抗体落下的复合物中。Ck2蛋白质免疫沉淀实验与Pcgf3和Pcgf5相互作用,尽管在质谱分析中有些蛋白质被漏掉了。

在几个蛋白质中,也出现在Pcgf3或Pcgf5的免疫沉淀物中。Tex10、Las1L和Pelp1与Senp3和Wdr18一起形成一个先前确定的复合物,进行了互相免疫沉淀实验, 以验证ES细胞中Pcgf3/5与5FMC复合物之间的相互作用。
在稳定表达带有FLAG标签的Tex10的ES细胞中进行FLAG-Tex10的免疫沉淀实验,允许Las1L、Pelp1和Pcgf3/5的共沉淀,这是Western blotting分析得到的。
在Tex10沉淀物中发现p300,显示Tex10与p300之间的相互作用来调控组蛋白乙酰化。在ES细胞中扩展了Pcgf3/5复合物的多单元组合,并且新的相互作用蛋白质,尤其是Tex10、Las1L和Pelp1,可能使这些复合物能够激活不被典型地认为是PcG功能的转录。

结论
Pcgf3/5在ES细胞中主要起到转录激活的作用,质谱分析进一步揭示了Pcgf3/5复合物中包含与转录激活有关的组分。
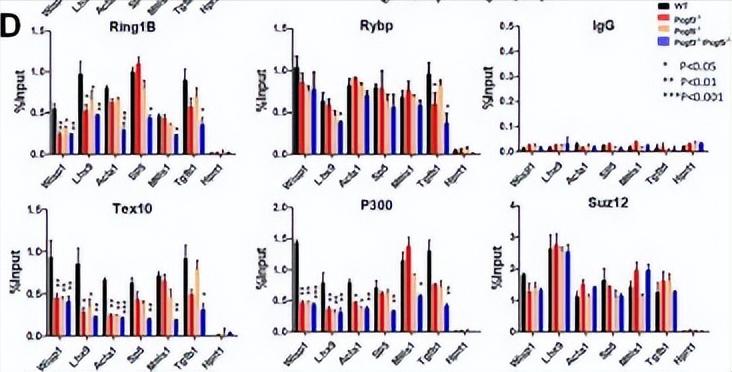
当Pcgf3/5对启动子上染色质特征的局部影响,对一组富集在中胚层分化中的目标基因进行了探测, 这些基因是通过全基因组RNA-seq分析选择的,它们的表达在Pcgf3/5缺失后下调。

Wisp1参与血管平滑肌细胞的迁移和增殖,Lhx9参与许多器官的正确发育。Acta1存在于肌肉组织中,是收缩装置的主要组成部分。Sp5在发育过程中表达动态,并与Brachyury发生遗传相互作用。
参考文献
[1] 张彩苹,细胞周期蛋白E在大鼠肺组织中的表达及其与气道重塑的关系,2010
[2] 宫晓庆,细胞周期蛋白依赖性激酶抑制因子3的研究进展,2020
[3] 朱海心,细胞周期素E在骨肉瘤和骨巨细胞中表达的实验研究,2002.
[4] 张宣,内组织p27蛋白、细胞周期素E和增殖细胞核抗原的表达及意义, 2003
[5] 吴晓明,浸润性导管组织中肽基脯氨酰顺反异构酶 1和细胞周期蛋白 D1的表达,2018