
介绍
广泛使用世界性和高产品种导致生物多样性严重丧失,威胁到许多地方种群的完整性。
然而,通常在边缘地区饲养的本地品种受到关注。
地方品种不仅是一种文化和遗传遗产,而且由于其强大的抗病性和对恶劣环境的适应性,也是一种基本的遗传资源,因此对自然资源的可持续利用至关重要。
这些品种需要仔细的基因组特征,以突出其遗传多样性,并为保护策略和育种计划提供有效手段。

在意大利,为避免这些遗传资源的完全灭绝,在20世纪80年*开代**始了当地猪品种的保护计划。
目前,全国猪饲养者协会(ANAS)已经正式认可并管理了八个意大利本土猪品种。其中,尼禄西西里亚猪品种代表了自希腊和迦太基统治以来西西里的遗产。
在19世纪,由于大量繁殖高产英国猪品种,以及大面积的森林砍伐减少了其繁殖面积,尼罗西西里亚的人口规模急剧减少。
2001年,尼禄西西里尼罗猪被正式认定为一个品种,然后,国家和地区的项目处理了它的保护和开发。

如今,《牧群书》中记录的动物约有6500头,其中约800头是母猪和公猪,其余的是用于生产新鲜肉类或加工肉类的动物。
根据品种标准,尼禄·西西利亚必须有黑色的被毛,背部-腰部区域有长长的刚毛,大部分黑色或部分白色的头部,前鼻轮廓直。
该品种一直在西西里岛东北部的半粗放型或粗放型系统中饲养,通过生态相容的做法开发边缘林地。
在这种情况下,该品种已经获得了对困难育种条件的特定适应,包括高抗病性、良好的繁殖能力和对劣质饲料的良好适应性。

在牲畜研究中,BeadChip阵列是最广泛使用的全基因组工具,可用于许多主题,包括基因组特征和结构、选择特征研究和QTL检测。
与其他物种类似,猪的全基因组分析已在当地、欧洲和全球层面进行,以估计遗传多样性、种群结构和分化模式的比率。
在本研究中,我们将Nero Siciliano与意大利其他地方和世界猪品种的SNP数据进行了比较,旨在了解遗传变异性、群体结构、纯合性(ROH)和杂合性(ROHet)模式的运行情况。
遗传特征和基因组结构的理解是制定保护计划和加强这种地方资源生产系统的基本步骤。

材料和方法
DNA采样
总共选择了来自80个不同农场的16头Nero Siciliano猪进行基因分型。
根据猪的表型特征和农民提供的信息选择猪,以尽可能多地收集无关的动物,并尽量避免混合或跨物种的动物。
在含有pH 8.0的EDTA作为抗凝剂的真空管中收集单个外周血(10 mL)。
授权人员根据指令2010/63 /EU执行采样程序,避免对动物造成任何痛苦。
使用商业Illustra血液基因组制备迷你旋转试剂盒从血液中提取DNA。

基因分型、质量控制和数据处理
所有动物均用Illumina PorcineSNP60 v2 BeadChip(Illumina)进行基因分型。
采用PLINK软件对NSIC = 80的Nero Siciliano猪进行PiHat试验。
根据品种内平均亲缘度,没有个体的将被移除。
然后将NSIC的SNP数据与从先前研究中检索到的基因型数据合并也使用PorcineSNP60 BeadChip获得。
最终数据集包括五个当地品种、四个世界性品种和一个野猪样本。
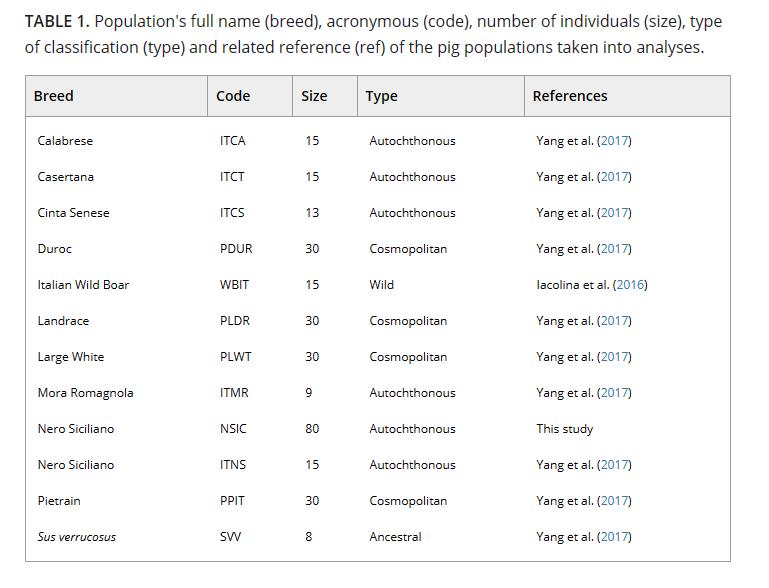
数据集组合如下(表1):卡拉布雷斯(ITCA = 15)、卡塞塔纳(ITCT = 15)、Cinta Senese (ITCS = 13)、Mora Romagnola (ITMR = 9)、意大利野猪(WBIT = 15)、杜洛克(PDUR = 30)、长白(PLDR = 30)、大白(PLWT = 30)、Pietrain (PPIT = 30)和另一个西西里尼罗(Nero Siciliano)样本(ITNS = 15)与实验样本(NSIC)进行比较。
软件Plink ver. 1.9用于执行过滤和质量控制。
每个SNP的染色体使用最新版本的猪基因组组装。
使用以下标准对基因型数据进行质量控制:次要等位基因频率≥0.02,SNP的基因型召唤率≥0.95和个体召唤率≥0.90,得到39 752例SNP和281头猪。
遗传多样性指数
Plink ver. 1.9用于估计种群内遗传多样性系数,即观察到和预期的杂合性,相对于每个群体的近交系数和次要等位基因频率。
当代有效种群大小使用NeEstimator ver. 2.1估计,然后再使用Waples和Do提出的连锁不平衡方法进行随机配对选项。
历史有效人口规模也通过默认设置选项使用gone软件计算。
遗传关系和种群结构
为了探究遗传关系和群体结构,利用Plink ver1.9中的——indep-pairwise 50 10 0.2函数对基因型数据进行筛选,去除高连锁不平衡(r2 > 0.2)的snp,从而生成了15 239个snp的修剪数据集。
Plink ver. 1.9还用于执行多维缩放(MDS)分析,基于个体之间的成对身份状态距离。
遗传结构由软件Admix ver.1.3.0估计,使用基于模型的聚类算法,范围从K = 2到K = 25并进行十倍交叉验证程序。
然后使用r包BITE ver 1.2.0008绘制估计的祖先矩阵,开源编程环境下的统计分析r 。
基于Arlequin推断的成对Reynolds遗传距离,构建了一个Neighbor-Net。3.5.2.2,并使用SplitsTree4ver. 4.14.8 绘制。
FST成对距离由Arlequin ver.3.5.2.2 计算。
最后,为了重建所有种群之间的基因组关系,并调查种群分裂和迁移事件模式的存在,使用TreeMix软件生成了一个最大似然树。
在本分析中,将疣状Sus样本(n = 8)作为外组。
执行了五次独立迭代,允许迁移事件的范围在1到10之间,而协方差矩阵使用每个块500个连续的snp来估计。
使用r包OptM中实现的Evanno方法评估最支持的迁移边缘数量。
结果
遗传多样性指数
表2报告了遗传多样性的汇总统计数据。
PLDR、PLWT和PPIT的H值最高E和 HO,其次是NSIC,而ITMR报告的值最低。
在ITCS, WBIT,尤其是ITMR中观察到高FIS值。
3个世界性品种和NSIC的FIS值较低,而ITNS、ITCA、ITCT和PDUR的FIS值处于中等水平。
本地品种中,ITMR的MAF值最低,NSIC最高;相似的MAF值在世界性品种中也有报道。
当代(cNe)有效种群规模报告于表2。
NSIC的cNe = 34.3。世界性品种的PPIT值最高,从30.4到50.7不等,意大利地方品种和WBIT的ITCT值最低,从2.5到20.1不等。
历史有效种群规模(hNe)在100代内的变化如图S1所示,呈对数趋势。
第1代的值与cNe相似,从第4*开代**始显著增加,在第19代趋于稳定。
在第31代,WBIT品种的曲线显著上升,在第71代,所有世界性品种和NSIC品种的曲线均显著上升。

品种结构
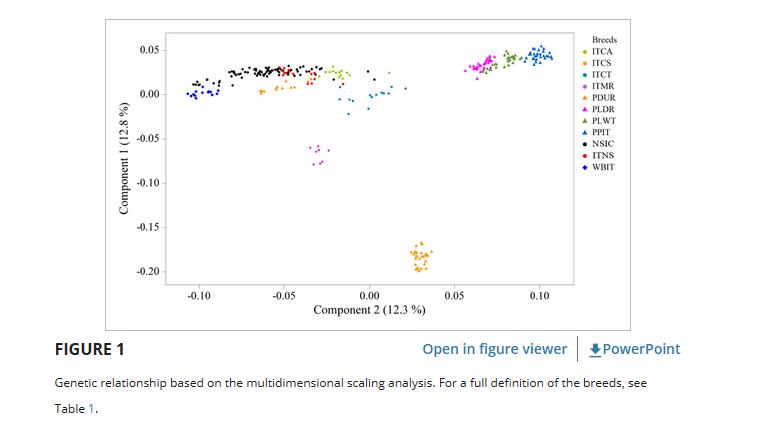
根据推断方差的前两个分量,MDS分析(图 1 )强调了世界性和本土品种之间的明显区别。
特别是,PLDR,PLWT和PPIT聚集成聚为一个内聚组,而PDUR与样品的其余部分分离。
组分C2显示出从左到右的变化梯度,几乎没有不连续性,首先识别WBIT和NSIC,然后是其他混合黑猪,接着是PDUR,最后是其他世界性品种的集群。
C1成分没有将WBIT与当地品种区分开来。
ITMR与其他没有相互分离的地方品种分离。
事实上,NSIC、ITCA和ITCS部分重叠,而NSIC和ITNS之间则如预期的那样完全重叠。在NSIC样本中,一组12只动物接近WBIT。
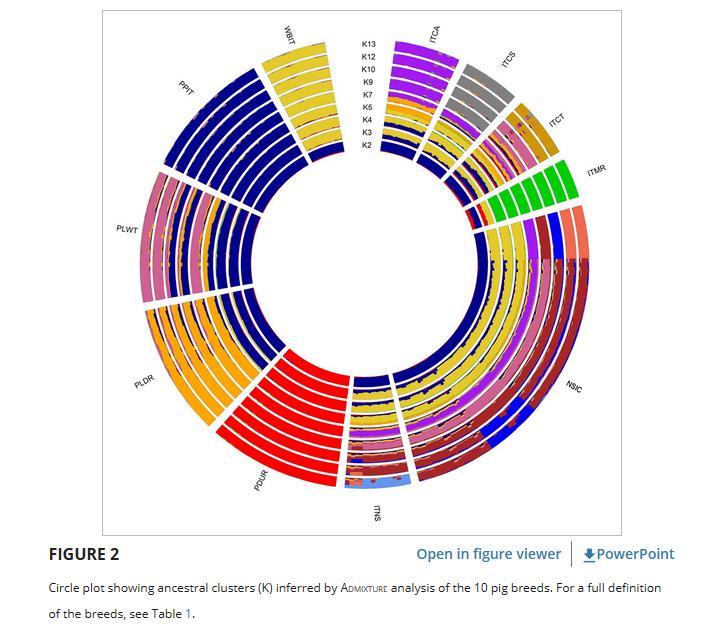
在混合分析中,推断聚类的交叉验证误差的分布(K = 25)表明K = 13是最有可能的祖先数量(图 S2 )。
圆图(图 2 )是指11个群体的基因组结构,显示了从K = 2到K = 13的最具代表性的祖先集群。
在K = 2时,种群的基因组模式通过将PDUR品种与其他种群分开来重现MDS第一组分的证据。
在K = 3时,区分了三个祖先簇,分别由PDUR,其余世界性品种和当地品种与WBIT一起代表。
在K = 4时,ITMR与其他意大利本地品种分开聚集。进一步的集群由PLDR和PPIT(K = 5)形成,其次是PLWT和WBIT(K = 7)。
NSIC和ITNS与其他当地品种共享共同的祖先,直到K = 7,而在K = 9时,它们形成了自己的集群。
此外,从K = 9到K = 13,NSIC显示出具有亚结构证据的异质遗传模式。
一般来说,PDUR、PPIT和PLDR报告了世界性品种中最高的基因组差异,而ITCA、ITCS,特别是ITMR强调了地方品种中最低的混合率。
尽管起源不同,但WBIT表现出高度的内部同质性,并对本土品种表现出基因组影响。
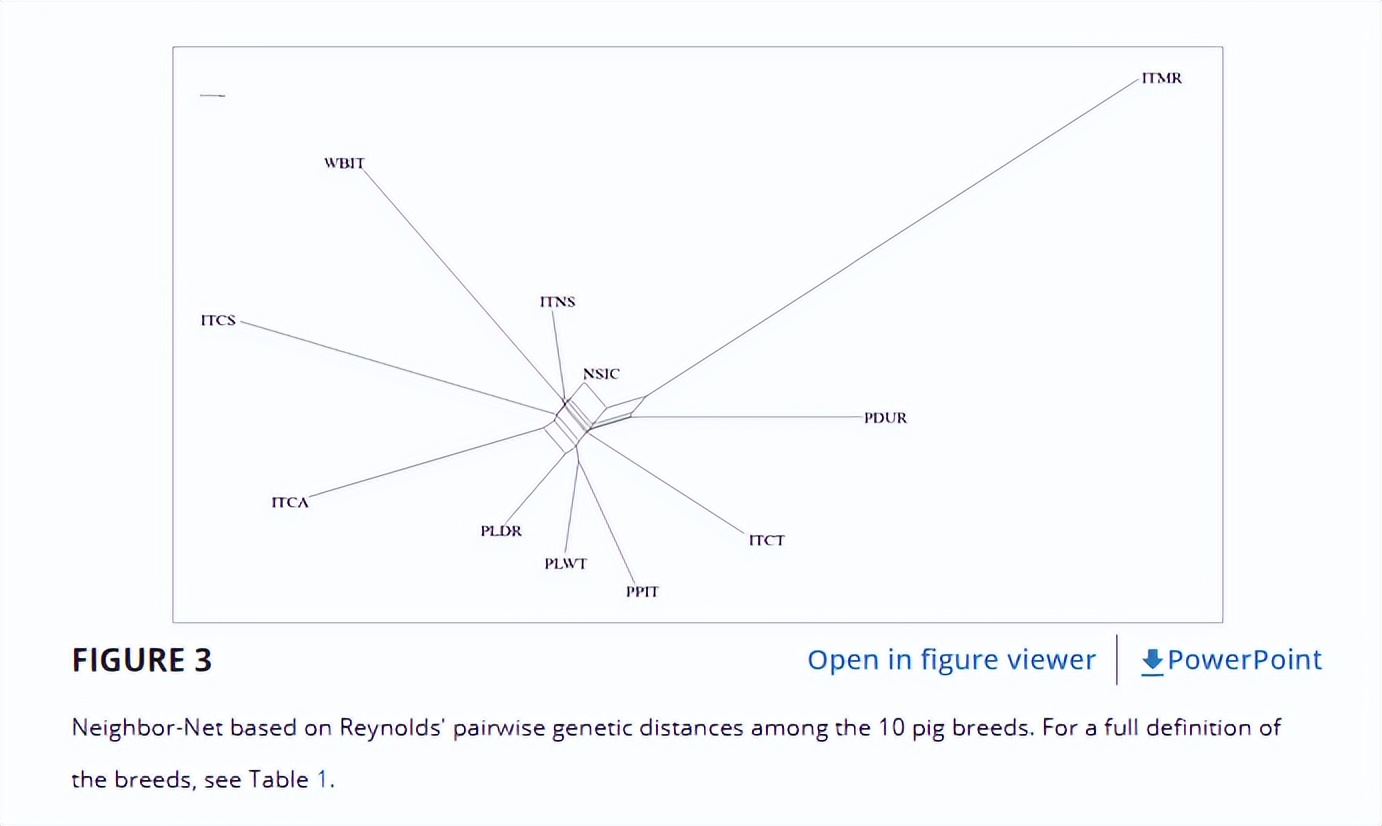
基于成对雷诺氏遗传距离的邻居网络(图3)显示了品种之间遗传关系的互补图景。
与MDS图一致,该图揭示了由NSIC,ITNS,WBIT,ITCA和ITCS组成的两个主要集群,另一侧由PLDR,PLWT,PPIT,PDUR,ITMR和ITCT组成。
在第一组中,WBIT更接近ITNS和NSIC,而不是其他品种。
在第二组中,ITMR与PDUR偏离了相同的系统发育节点,并突出显示了网络的最长分支;ITCT处于PLDR,PLWT和PPIT与ITMR-PDUR组形成的集群之间的中间位置。
FST成对距离(表S1)突出了ITMR与WBIT(0.453),与ITCS(0.451)和与ITCA(0.422)比较的最高值,而NSIC与ITNS之间的最低值为0.048。
NSIC与ITCT(0.116)、PLDR(0.128)和PLWT(0.130)的平均距离最低(0.132),相对较低。
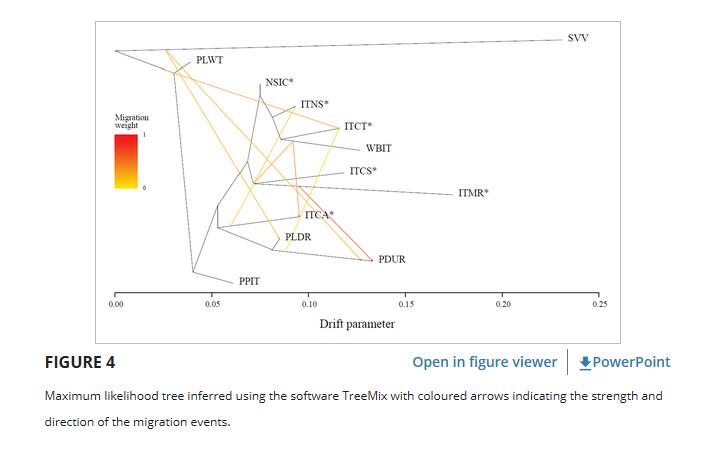
TreeMix树状图(图 4 )基本上证实了先前的分析,表明除ITCA外,所有意大利本地品种都聚集在一起。
然而,ITCA是意大利品种处于基础位置的小组的一部分,其中还包括PDUR和PLDR品种。
意大利野猪属于意大利当地品种,而其他世界性品种则处于基础位置。
Evanno 方法指出,最受支持的迁移事件数为8个。
从外群到PLDR和PDUR显示了两个迁移边缘,另一个迁移事件从分支的底部突出显示,包括所有品种到ITCT。
WBIT显示出向ITCA和节点的两个迁移边缘。
从ITMR到PDUR的迁移权重估计最强,而从PDUR到ITCT检测到的最弱信号。
NSIC与ITNS,ITCT和WBIT一起分支,没有报告迁移事件,而从ITCA的基础到ITNS的迁移边缘较弱。
结论
本研究报道了对经济上重要品种如尼禄西西里亚猪的基因组结构的深入调查。遗传多样性指数表现为中等水平的变异。
几种统计方法的结果表明Nero Siciliano具有明确的遗传背景,并强调了其与意大利地方品种和野猪的遗传关系。
这可能是由于尼禄西西里亚猪与野猪自然杂交的半粗放繁殖系统。
该地方品种的遗传异质性表现在其相对较低的近交系,与世界性品种相当。

分析样品的内部亚结构可能决定了纯合区域的适度固定和杂合区域的几乎零固定。
此外,在西西里亚诺发现的纯合子岛强调了它与野猪和其他意大利品种的关系,并强调了与生产性状相关的基因的固定,这可能是过去与商业品种联系的痕迹。
对杂合性丰富区域的调查突出了世界性猪品种中几个与免疫相关的热点,表明充分的配种计划不会损害它们的适合度反应。
这项研究证实了历史和科学报告先前对尼禄西西里亚人的描述。此外,它强调需要适当的配对计划,使其基因组结构同质,从而提高品种和产量。
