
在阅读此文前,麻烦您点击一下“关注”,既方便您进行讨论和分享,又能给您带来不一样的参与感,感谢支持!
文|嫋嫋
编辑|嫋嫋
前言
天然棕黑色真黑色素为动物提供了结构着色,并能有效阻挡电离辐射和抗真菌药物。这些功能使得它们在仿生材料设计方面具有吸引力,用于增强水凝胶材料的添加剂以及用于土壤修复的自由基清除剂。

然而, 由于这些复杂聚合物组分的不溶性、异质性和无定形特性, 导致真黑素分子结构和相关功能的分子决定因素和结构特征仍然不确定。
儿茶酚胺前体改变黑色素在真菌细胞或无细胞条件下的结构
改变黑色化真菌细胞的支架组织和膜脂含量,使真菌细胞优先使用l-DOPA前体进行黑色化,无细胞和真菌真黑色素的大分子碳基和氮基结构包括吲哚、吡咯、吲哚醌和根据反应时间形成的开链结构单元。

儿茶酚胺前体的可用性对于影响色素形成的效率、真黑素分子结构及其在真菌系统中的基本支架起着重要作用。
黑色素在许多方面都具有多种功能, 赋予动物结构着色、阻挡电离辐射、抗真菌药物和能量转换等。这些功能使得黑色素聚合物复合材料和结构相关的聚多巴胺成为仿生材料设计的有吸引力的平台。

尽管这些材料具有实际重要性和多功能性,磁共振波谱、稳定同位素富集、计算建模和受控化学反应等方法在理解真黑素结构方面取得了进展。
真黑素的分层沉积依赖于多糖细胞壁支架,无细胞和基于新型隐球菌合成的真黑素显示出常见的基于吲哚的芳香核。不同比例的儿茶酚胺前体,可以影响聚多巴胺合成及相关超分子结构的光学和导电性能。
但黑色素合成的具体底物尚不清楚, 因为脑组织富含儿茶酚胺 ,在不同前体的黑色化对于深入了解脑组织中黑色素合成的可能途径非常重要。
儿茶酚胺前体是否控制无细胞合成黑色素产物的结果、特定前体的可用性是否影响黑化真菌细胞壁支架的多糖和脂质组织、新型隐球菌黑化的首选生物合成前体细胞是哪些,以及在无细胞系统中,哪些黑色素构建块和大分子组织可以被识别为反应时间的函数。

前体身份改变无细胞和真菌黑化中的色素结构
新型隐球菌真菌细胞提供各种儿茶酚胺前体后,形成了结构多样的黑化细胞壁材料(“幽灵”)。这些材料在生物合成和大分子结构方面的差异,可能包括细胞壁支架对不同前体黑色素沉积的能力、各个单体的聚合途径以及这两种原理的组合。
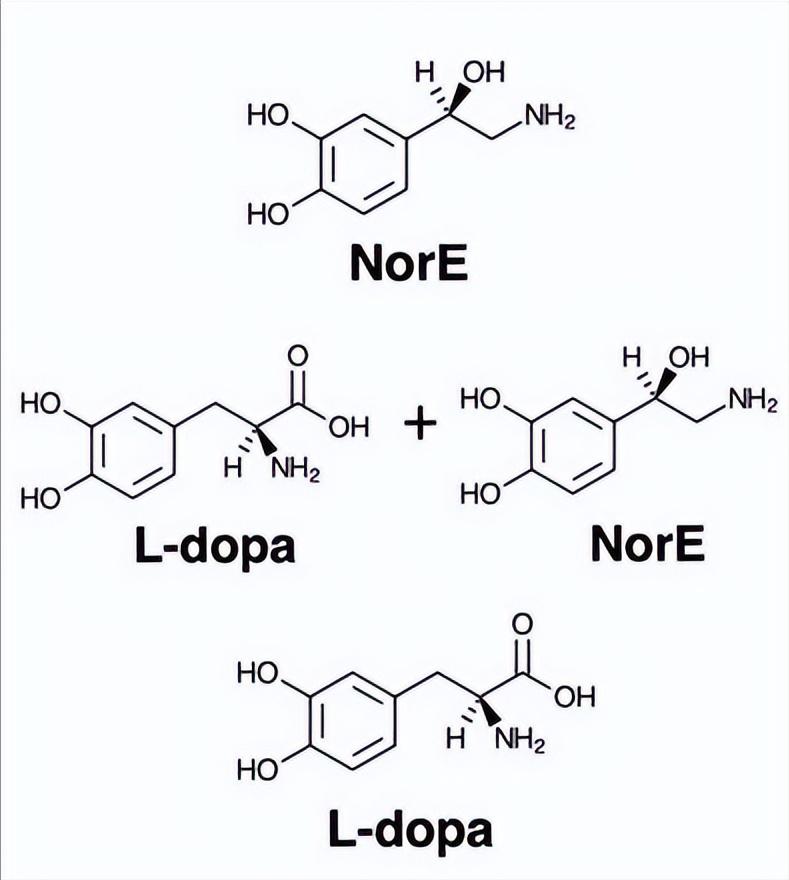
在该真菌系统中拥有几种儿茶酚胺前体(l-DOPA、甲基-l-DOPA、去甲肾上腺素(NorE)和(肾上腺素), 起始材料及其各自的不溶性固体产物的固态13C NMR光谱指纹。

与聚合物组装体形成相结合后,生物合成颜料的13C CPMAS光谱显示出改变的共振位置和线宽。
尽管新型隐球菌真菌细胞和无细胞系统中形成的无定形黑色素“幽灵”通常具有广泛的共振,但使仍然可以识别由羧酸盐或酰胺结构、芳烃和烯烃、多糖细胞壁的烷氧基和烷基链的NMR信号。

尽管儿茶酚胺前体均具有典型的尖锐,但自聚合产生的无定形聚合物和 C. neoformans均具有典型的结晶固体特征, 黑色素“幽灵”在其光谱的芳香族和脂肪族区域显示出更广泛的特征。
通过任一合成方法形成的固体芳香族黑色素颜料和黑化脂肪族细胞壁,都具有无定形特征,无论是否存在酶或支持支架。

颜料非常宽的芳香族光谱包络可能是由于共振重叠(化学位移异质性),而不是自由基导致的自旋弛豫效应,因为许多NMR共振在选择性13C标记材料中得到了更好的解决。
芳香族共振的广泛出现可以反映稍微不同的DHI、DHICA和其他可能的结构构件,不同单元间的共价连接,以及连续颜料层的异质排列堆叠。

每种纯化的黑色素显示出明显的脂肪族信号,这些特征与各自儿茶酚胺前体的不同。虽然真菌黑色素制剂的 30 ppm和72 ppm 共振可能源自黑色素颗粒中的细胞膜和细胞壁成分,但这种解释不能解释相应无细胞制剂光谱中相同或相似的特征。
对新型隐球菌黑色素提出了另一种假设,该假设涵盖了两种制备方法与聚多巴胺类似,将脂肪族结构视为保持未环化或部分聚合的前体部分,同时形成共价结合的聚合物。

这种不完全形成或“发展”芳香族黑色素结构的提议在重新审视黑色素生物聚合物的分子结构时具有重要意义,其中包括吲哚、吡咯、开链和新提出的结构。
在儿茶酚胺前体中, l-多巴和甲基-l-多巴产生了最强烈的黑色素沉积, 由它们的吲哚基结构产生的芳香核磁共振信号证明,而肾上腺素在无细胞或真菌反应中基本上不形成吲哚样色素。

这与之前的新型隐球菌制剂一致,其中来自l-DOPA、甲基-DOPA、去甲肾上腺素和肾上腺素底物的黑色素产物的 产率分别为47%、61%、27%和2%。
尽管在13C光谱中未检测到色素的肾上腺素衍生物,其细胞壁部分黑化,因为未黑化的细胞无法在分离鬼影的严酷程序中存活下来。

通过一维自然丰度光谱指纹只能对无细胞和真菌制剂进行定性比较,与使用二维13C-15N固态NMR相对应的结果一致,并支持了芳香核的共性证据。
前体身份改变真菌黑色素鬼细胞支架的结构顺序和脂质含量
当l-DOPA和甲基-l-DOPA产生的细胞黑化,支持了含有DHICA和DHI的芳香色素的形成需要稳定的环状多巴色素中间体结构的主张,真菌黑色素前体结构中羟基取代的苯或吲哚环的要求也强化。

新型隐球菌可以从血清素和4-羟基吲哚形成芳香色素,在较小程度上从5-羟基色氨酸和5-羟基吲哚形成芳香色素, 但不能从色氨酸本身形成芳香色素。
除了儿茶酚胺前体在黑色素合成中的作用外,还着重了解潜在的黑化细胞壁结构对于新型隐球菌系统的重要性。已知色素沉积会影响颗粒形成、细胞孔隙率和耐药性。新型隐球菌真菌系统中,葡萄糖衍生的脂肪族支架是在黑色素形成之前形成的。

黑化细胞内紧密堆积的颗粒层的逐渐发展提出了一个问题:这些层是否只是简单地彼此交联,或者通过由色素、蛋白质、多糖和/或脂质组成的支架保持在一起。黑化细胞壁必须具有足够的可塑性,以允许细胞出芽和形态转变,例如菌丝形成。
黑色素鬼中保留的细胞壁支架是否与色素来源的儿茶酚胺在分子组成、顺序或组织上有所不同,使用固态13C NMR来检查葡萄糖一起生长的新型隐球菌真菌细胞的细胞壁组成。通过比较使用不同前体(l-DOPA或NorE)生产的黑色素细胞的光谱。

NorE黑色素细胞的细胞壁结构比l-DOPA黑色素细胞更有序, 含有更多的几丁质多糖和膜脂质成分。这可能是导致NorE黑色素沉积较少的一个原因。
总结
除了前体结构,对黑色素形成过程中的细胞壁结构,对于了解黑色素合成的细节和细胞的生理特性至关重要。

不同儿茶酚胺前体对新型隐球菌细胞进行了是否优选特定前体或是否可以形成具有独特大分子结构的色素。当使用l-DOPA和NorE儿茶酚胺的1:1混合物作为黑化底物时, 所得的13C NMR谱与l-DOPA真菌黑色素非常相似, 表明l-DOPA是新型隐球菌细胞中的首选底物。
DARR也支持了这种观点,显示l-DOPA黑色素与NorE黑色素相比具有更宽的交叉峰和较弱的对角峰,这表明l-DOPA黑色素含有足够的脂质或有序性的无定形结构,以允许偶极串扰。

此外,EPR光谱结果表明,新型隐球菌细胞中使用l-DOPA或1:1混合物黑化的黑色素鬼影在信号强度和线宽方面与NorE黑色素相似。
这些结果支持一种机制模型,即去甲肾上腺素与l-DOPA一起在聚合过程中不会形成吲哚基色素,但新型隐球菌真菌能够单独利用每种底物来实现保护性细胞壁黑化。

另外,与使用儿茶酚胺黑色化的脂肪族成分相比,使用葡萄糖衍生的支架在光谱中显示出优异的分辨率。因此, 肾上腺素与l-DOPA一起产生了色素沉积的标志, 但使脂肪族支撑支架缺乏脂质且组织有序。
这些可以合理化为,如果两个前体发生缔合或共价结合以暴露亲水性-OH基团,从而促进颜料锚定到多糖支架上,这将导致脂质在鬼分离方案期间非常容易被提取。

在一组富含核磁共振活性同位素的现成儿茶酚胺前体中,黑化过程构建的含碳和氮的异核分子框架。
在细胞培养过程中,l-DOPA或多巴胺前体会逐渐产生增加的色素沉积量。合成的多巴胺聚合物具有表示吲哚、吡咯和未环化儿茶酚胺结构单元的碳和氮部分,并且真菌和合成黑色素中可以识别出几种结构类型的类似近端碳-碳和碳-氮对。

在使用二维双共振固态核磁共振策略,能够限制一系列真黑素色素的可能结构, 并获得对黑色化发展进程的新分子。
通过2D 13C-15N Z 过滤转移回波双共振(ZF-TEDOR)实验可以关联NMR信号,揭示位于高达5Å距离范围内的近端碳氮对各自的化学环境,避免了可能产生的偶极相移效应。

研究比较了在1.5天时形成的合成L-酪氨酸黑色素的短TEDOR重耦合时间获得的二维光谱与成熟3.0天的光谱。
通过与增强灵敏度的动态核极化(DNP)相结合,可以观察到固相黑色素聚合物合成的发展过程。实验识别出13个C-15N对,它们之间的距离在约1.5Å范围内,可能代表定义聚合黑色素结构的直接键合的异核。
参考文献
[1] 戴均贵,朱蔚华,吴蕴祺,等.前体及真菌诱导子对银杏悬浮培养细胞产生银杏内酯B的影响.药学学报.2000.02.017
[2] 李祝,郁建平,梁宗琦,等.前体物质与真菌激发子对虫草菌素产量的影响.食品科学.2008.06.057
[3] 朱蔚华.前体及真菌诱导子对银杏悬浮培养细胞产生银杏内酯B的影响.药学学报.2023.08.02
[4] M·华德,Y·罗,F·O·本德苏,等.在不存在诱导底物下丝状真菌细胞中的蛋白产生.2023.08.02
[5] 杨安全,王菁,沈玥琦,等.珍珠提取物对黑色素细胞酪氨酸酶活性和黑色素合成的影响.药物生物技术,2018-04-007