
点点关注不迷路,更多精品等着你!
通路,信号通路是也,英文名称Signaling pathway/Signal transduction。它的特点就在于可仅凭一人之力就撑起了分子机制的整个天下。通路的本质在于分子变量,是由一组已知的、且确定上下游信号传导关系的明星分子所组成的。显然,每一个通路都有代表性的标(Biomarkers),因而只要弄清楚它们就可以见微知著了。
1
信号通路基本常识
生物界的信号有三类:物理信号、化学信号和生物信号,而人体自身的信号转导主要依赖的是生物信号,比如内分泌系统的激素、神经系统的神经递质以及通过旁分泌或自分泌产生的细胞因子等。而信号作用于细胞后会有5种常见的结局:促进代谢,细胞分裂,细胞分化,细胞死亡或者其他——激活某一种特定的细胞功能。
通常信号通路包含三个基本的构成元件:
1)配体(ligand)和受体(receptor)。当配体特异性地结合到细胞膜或细胞内的受体后,可通过构象改变以及蛋白磷酸化修饰对下游一系列蛋白进行激活或抑制调节,将外界信号级联放大,最终产生综合性的细胞应答反应。
2)蛋白激酶(kinase)。它可将ATP的磷酸基转移到底物某个蛋白的特定氨基酸残基上去,从而快速改变下游蛋白的构象,是常规信号通路中传递信息的最主要的蛋白类型。激酶中最主要的两类是酪氨酸激酶(PTK)和丝氨酸/苏氨酸激酶(STK)。
3)转录因子(transcription factor)。它是一类DNA结合蛋白,可参与调控基因转录过程,成为了胞内的第三信使。而第一信使是胞外的配体,第二信使是配体受体结合后激活的胞内信号分子,如环磷腺苷(cAMP)、环磷鸟苷(cGMP)、Ca2+等,有助于信号向胞内进行传递。
2
9条炙手可热的信号通路
现阶段,科研中研究最为火热的信号通路有9种,分别是NF-κB、PI3K/AKT/mTOR、MAPK、JAK-STAT、TGF-β、Wnt、Notch、Hedgehog和Hippo通路。跨越通路学习的初级阶段时,无需深挖各种细节,只需把握路通中最关键的几个biomarker即可。
1. NF-κB信号通路
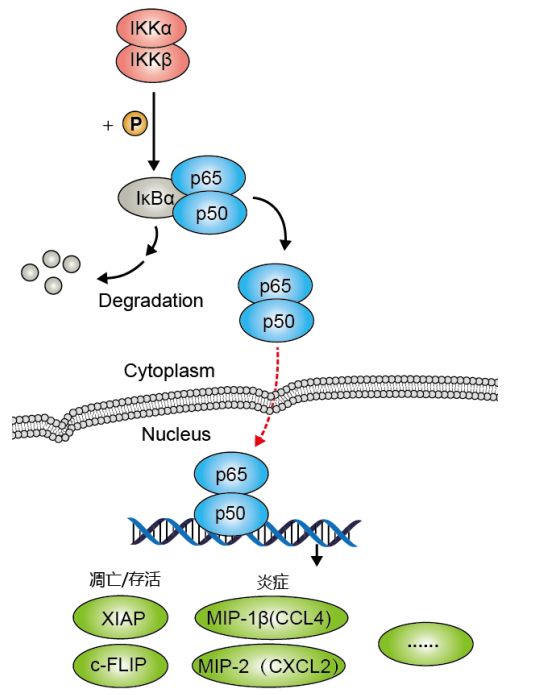
(图片截取于优秀学员Ewen的整理笔记)
NF-κB蛋白通常会由p65和p50形成同源/异源二聚体,在胞质中因与抑制蛋白IkB结合形成了三聚体复合物而处于失活状态。当上游信号因子TNF结合到细胞膜表面受体后,受体构象改变并将信号传递给IKK激酶(IkB kinase),进而使IkB蛋白磷酸化并将其从三聚体中解离出来。
随后NF-κB二聚体暴露出核定位序列(NLS),迅速从细胞质进入细胞核内,与核内DNA上的特异序列相结合,促进相关基因的转录,比如CyclinD1、c-Myc、MMP-9、VEGF等。因而该通路的持续激活会导致细胞生长失控。
NF-κB通路与癌症有着密不可分的关系。NF-κB能转化细胞的证据最初来源于人们从一种禽类逆转录病毒中发现的v-Rel基因,此基因编码的蛋白质有高度的致癌性,能使鸡产生恶性肿瘤。后期研究发现,v-Rel编码的蛋白属于NF-κB的Rel亚家族,而且其他陆续发现的几种致癌病毒也是通过NF-κB的激活来致癌的。
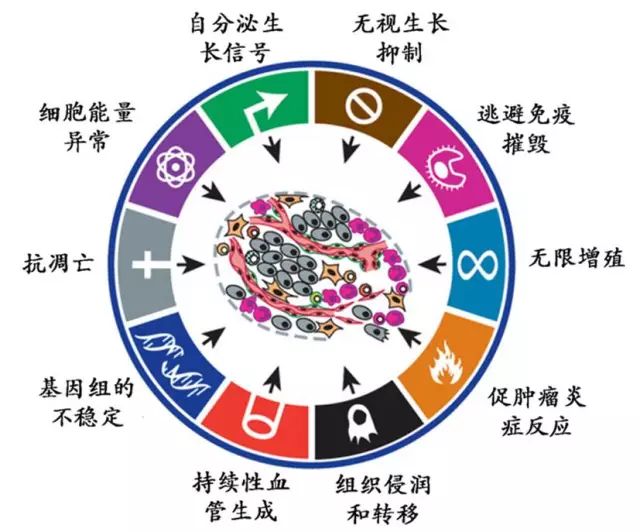
来源:D. Hanahan, R.A. Weinberg, Hallmarks of cancer: the nextgeneration, Cell, 144 (2011) 646-674.
这张图总结了癌症细胞10大标志性特征,而NF-κB通路几乎是里面所有的帮凶。
周期蛋白D1、周期蛋白依赖激酶Cdk2、c-myc等基因转录受NF-κB的调节。在乳腺癌、结肠癌、淋巴癌等癌症中,NF-κB的持续性激活导致与细胞周期相关的蛋白活性异常。
其次,持续性激活的NF-κB能提高一些促进肿瘤生长的细胞因子水平,如IL-1β(急性白血病生长因子);TNF(恶性淋巴肉芽肿瘤、T细胞淋巴癌、神经胶质瘤生长因子);IL-6(多发性骨髓瘤的生长因子)。
肿瘤发展的两个重要阶段是组织侵润和转移,这两个过程也是由NF-κB依赖的基因所调节的,包括基质金属蛋白酶、尿激酶纤维蛋白溶酶元激活剂以及白细胞介素IL-8。此外,肿瘤细胞的生长和组织侵润需要不断有新血管的形成,而介导新血管生成的相关蛋白的基因受NF-κB的调节。其中就有血管生成因子家族中最重要的成员——血管内皮细胞生长因子(VEGF),持续性激活的NF-κB能增强VEGF基因的转录。
最后,NF-κB的持续性激活也能导致细胞的抗凋亡,抵抗化疗药物的诱导凋亡作用。其主要机理是NF-κB可以大量抑制细胞凋亡相关基因的转录,如细胞凋亡蛋白抑制物c-IAP1/c-IAP2、肿瘤坏死因子受体结合因子TRAF1/TRAF2和白介素转换酶抑制蛋白cFLIP,以及锌指蛋白A20。
以上介绍的几种NF-κB介导的肿瘤形成机制能够相互协调,在不同信号传递水平上相互作用,共同促进肿瘤的发生与发展。目前针对NF-κB的肿瘤靶向治疗是癌症治疗的热点之一,其主要思路是抑制NF-κB的活性;抑制IκBα蛋白的磷酸化,避免其被泛素化降解;抑制NF-κB的DNA结合活性等。
然而,需要强调的是,NF-κB同时也具有介导免疫应答的正常生理功能。机体受感染后的炎症反应需要启动NF-κB信号通路,转录一些细胞因子来介导免疫应答,以清除入侵的病原菌。对NF-κB的抑制在缓解肿瘤发生发展的同时,也将抑制正常的免疫应答,所以如何平衡这两者的关系,也是必须考虑的重点。
2. PI3K/AKT/mTOR信号通路
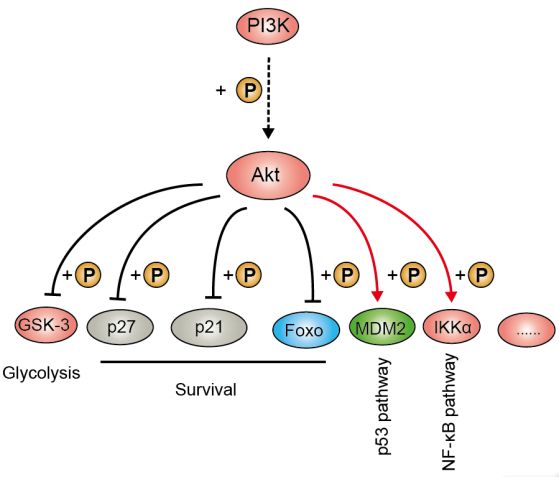
(图片截取于优秀学员Ewen的整理笔记)
PI3K/Akt的Biomarker是PI3K和Akt的总蛋白以及磷酸化蛋白。PI3K(磷脂酰肌醇激酶)是由调节亚基p85和催化亚基p110构成二聚体。当它与生长因子受体(如EGFR)结合后,可改变Akt的蛋白结构并使其活化,并以磷酸化作用激活或抑制下游一系列底物如凋亡相关蛋白Bad、Caspase9活性,从而调节细胞的增殖、分化、凋亡以及迁移等表型。另外,Akt也可激活IKK,与NF-kB通路存在cross-talk。
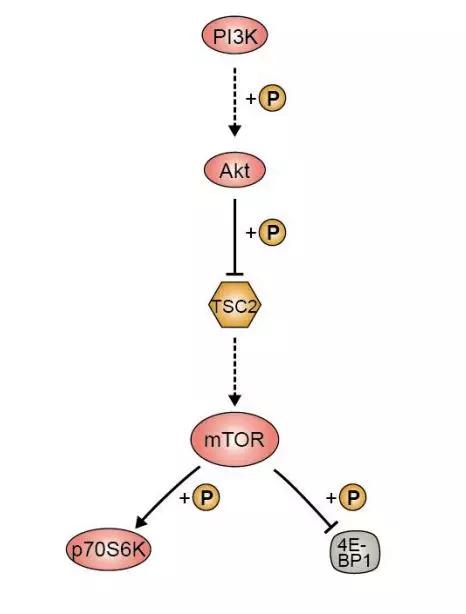
(图片截取于优秀学员Ewen的整理笔记)
PI3K/Akt下游靶点是哺乳动物雷帕霉素靶蛋白(mTOR),而mTOR的下游转录因子则包括了HIF1α、c-Myc、FoxO等明星分子。值得注意的是,抑癌蛋白PTEN作为磷酸酶,可使Akt去磷酸化而减少活化,可阻止所有由Akt调控的下游信号传导事件,是PI3K的负向调节因子。
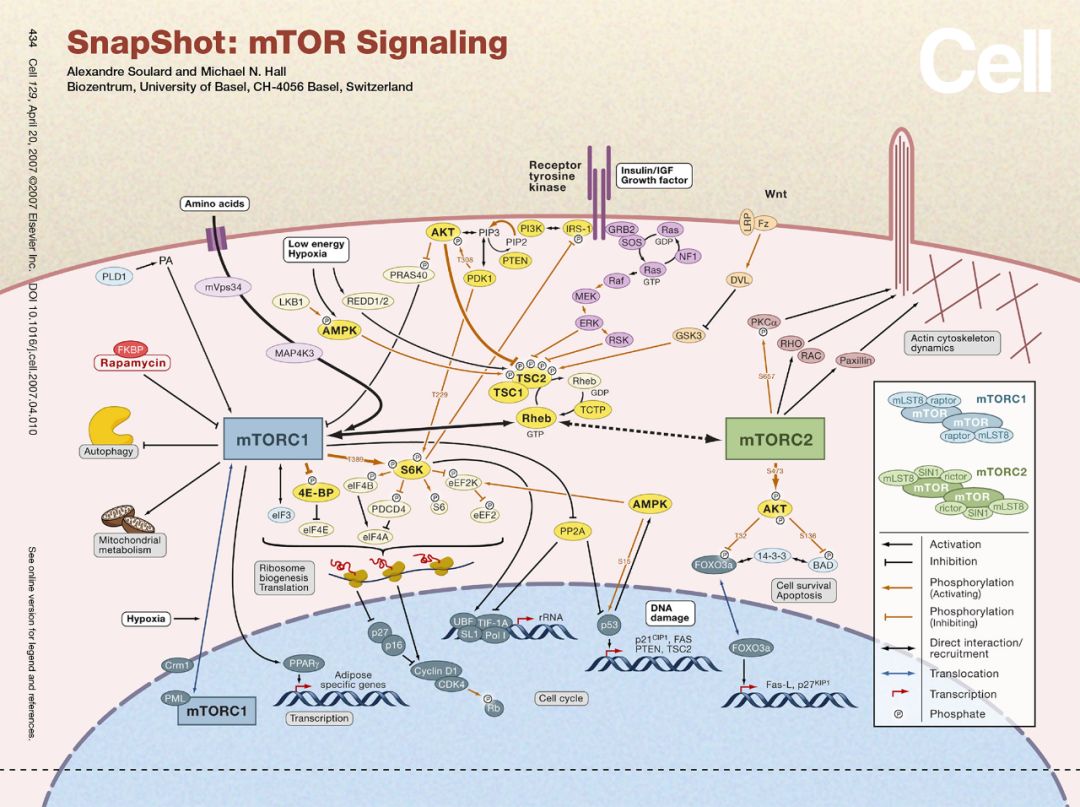
能量的代谢通常都伴随着物质的代谢,物质循环也伴随着能量的产生与利用,这样的过程需要根据细胞与机体的物质和能量的需求状态进行精细的调节以维持生命活动的稳定。而明星分子mTOR则是调节生长与代谢的这样的一个枢纽分子,那它具体是怎么参与代谢调控的呢?
2.1、对胰岛素和胰岛素样生长因子的响应
PI3K-AKT-mTOR是经典的响应胰岛素信号的通路。当进食后,被分解的葡萄糖进入血液促进胰岛素的释放,胰岛素作为响应营养富余的信号,会指导细胞进行吸收利用这些营养。
Insulin首先结合细胞表面受体通过IRS1激活PI3K-AKT通路,AKT直接促进Glucose的吸收,同时通过AKT-TSC1/2-RheB-mTORC1激活mTORC1的活性,mTORC1进一步指导合成利用glucose进行生物合成相关的酶进行营养的储存。RheB是一种小G蛋白,小G蛋白激活后可直接调控mTORC1,介导的激素类即细胞表面信号到mTORC1,这是众多mTORC1调节方式中的一种方式。
这一过程既包括吸收也包括吸收后的利用调节,这样一条龙的调节方式,通常被很多科研工作者理解为mTORC1被激活后,既可以合成糖原、可以促进翻译,也可以促进生长。
这样笼统的理解显然是有失偏颇的,也是我们经常对文献理解的一个误区。实际上Insulin激活mTORC1大部分情况下是和糖代谢相关,而IGF(胰岛素样生长因子)激活mTORC1才主要涉及细胞的生长相关。mTORC1激活后分别调控的底物理论上是有差别的,非常具体的调控与底物信息等仍有待进一步研究。
2.2、对氨基酸-营养物质的响应
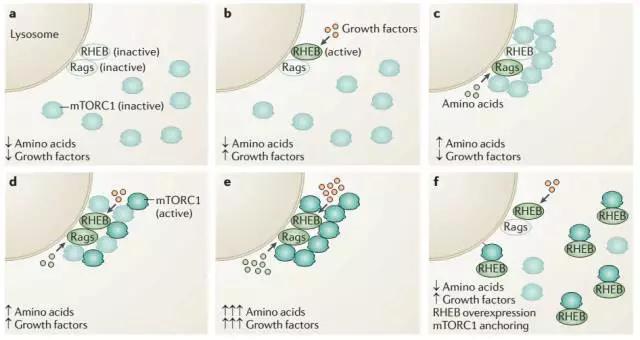
三大营养物质其中糖类,前面已经提到主要通过Insulin与细胞表面受体间接调节mTORC1的活性。而氨基酸被吸收后则可以直接激活mTORC1的活性。Sabatini发现mTORC1的激活主要有2步:
首先在氨基酸存在的情况下,小G蛋白Rags被激活,结合mTOR协助其定位于溶酶体膜表面。然后定位于膜表面的小G蛋白Rheb蛋白被来自生长因子的信号激活后才可以激活mTOR的活性。这里的由氨基酸直接激活mTOR的调节模式虽然不经由来源于细胞表面的信号,也是由小G蛋白Rags所介导。
既然在growth factor和氨基酸都存在的情况下mTOR才可以被完全激活,那也就是在物质和能量都富余的情况下才具备活化mTOR的条件。这一点正说明了mTOR作为代谢调节的枢纽具有信息整合的作用。试想一下,如果glucose富余而氨基酸短缺的情况下,细胞拿什么材料去合成,来完成生长呢?
所以这个时候glucose更多的是用来转化成氨基酸而非能量储存,所以mTOR未被激活。相反在氨基酸富余而glucose不足的情况下,合成所需的能量补给不足,氨基酸这个时候更多的可能会被用来分解而提供细胞活动所需能量,所以mTOR也必然不能被激活。
这正看出mTOR的调节模式也反映了细胞代谢平衡机智的策略。比较有意思的是,两条线路都是经由小G蛋白的信号,那么细胞的代谢稳态比如氨基酸和糖类之间的生化转化的信号如何感知呢?是否小G蛋白参与其中起到重要的作用呢?引人遐想。
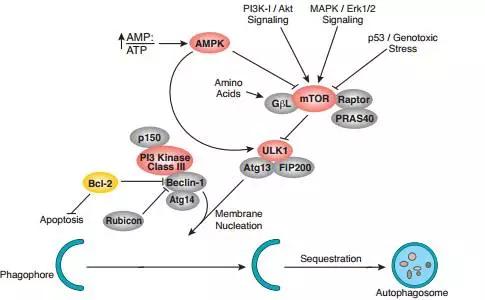
2.3、对自噬的影响
mTOR对自噬的调节本质上也是生长和代谢之间的调节。这其中有营养因素,也有能量因素。自噬是当细胞的营养物质或能量不足时,细胞为了维持自己的基本生存需要,就会通过溶酶体降解一些相对次要的蛋白,以及一些相对多余的细胞器,来供给物质和能量。
除了能量的主要载体ATP相对不足,即AMP/ATP上升,营养供应不足,mTORC1的活性也会不足,于是mTORC1释放了对自噬起始调节的抑制。另一方面来自PI3K-AKT,MAPK cascades信号对mTORC1的调控也体现了生长调节与代谢之间的联系。此外,P53/基因组的稳定性也可直接调节mTORC1的活性,也增加了这方面的证据。
2.4、对肿瘤治疗的影响
既然mTOR对代谢有这么大的影响,众所周知,肿瘤就是个能耗大户,那抑制mTOR通路是不是就能影响肿瘤的生长呢?
还真有这种研究,雷帕霉素就是经典的mTOR的*制剂抑**,能够同时抑制住通路中两个大块头mTORC1和mTORC2的活性。那是不是只要用它就能很好的限制住肿瘤呢?
没那么简单,下面的一个负反馈的图就很好的说明了这个原因。
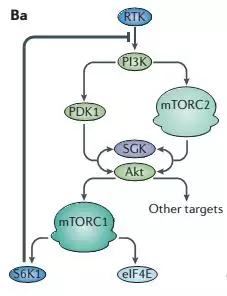
当mTOR被长期抑制后,S6K1的活性也会降低,S6K1对RTK的反馈抑制回路就会被解除,那么,将会有更多的补偿回路来补偿AKT和AGKs这些促进生存的因子,此外S6K1还可以抑制MEK-ERK信号通路,同样雷帕霉素可以解除对MEK-ERK的抑制,抑制临床上表现为雷帕霉素抵抗性。
有研究表明,雷帕霉素并非完全抑制了mTORC1的全部活性,在不同的底物可能各有差别。对于4E-BP1只是短暂或部分抑制其磷酸化,当S6K1-IRS1的负反馈回路使得PI3K-AKT的激活,最终可能还会导致4E-BP1的过磷酸化以及mTORC1其他促进肿瘤的功能。
科学家试图设计mTOR的*制剂抑**以期对抗肿瘤。竞争ATP结合的*制剂抑**Torin 1和PP242都有很好的临床疗效,但仍然无法回避S6K1-IRS1的反馈回路。再想下去,如果保持mTORC1的活性保持对AKT活性负反馈的完整性,而只抑制mTORC2的活性。
这样就可以抑制AKT的Ser473的磷酸化抑制其活性,然而后续的研究却发现,虽然AKT的Ser473的磷酸化被抑制,理论上Thr308的磷酸化不会被引发,实际的情况却是在此背景下Thr308位可能通过其他的路径得以补偿,mTORC2的抑制对AKT活性的影响恐怕只会影响到部分底物如FOXO1和FOXO3。
而最近几年的策略集中在对mTOR和PI3K双重*制剂抑**,这两种蛋白激酶都属于PI3K蛋白激酶家族。从原理上说,它不仅抑制了mTOR的活性,也直接抑制了AKT的活性,所以很好的规避了S6K1-IRS1的反馈回路。双重*制剂抑**PI-103也展示了良好的杀死淋巴癌的效果,但是这样的双重*制剂抑**对正常的淋巴组织却一样有毒副作用。针对mTOR的肿瘤治疗方案依然任重道远。
对mTOR靶向治疗肿瘤的尝试也很好的证实肿瘤是一个种调动了整个系统的系统性疾病,这大概是肿瘤难以对付的最重要的原因。对于简单的病因的肿瘤,治疗则相对较为容易。
对于复杂的病因引起的肿瘤,可能最初也是由单因素引起的细胞内部对生长增殖自我约束机制的破坏,而简单点的破坏可能会被自我修复,也可能引起连锁的正反馈的约束的逃逸,引发肿瘤。这好比细胞社会中,如果机体受到的外界因素的伤害不能抵御时,那么细胞个体便会不依赖整体协作而作为个体而努力存活下去。
3. MAPK信号通路
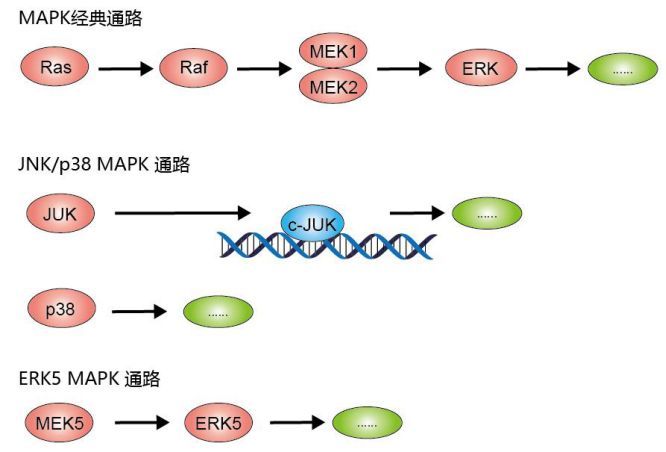
(图片截取于优秀学员Ewen的整理笔记)
MAPK(丝裂原活化蛋白激酶)通路有三级的信号传递过程:MAPK,MAPK激酶(MEK或MKK)以及MAPK激酶的激酶(MEKK或MKKK)。这三种激酶能依次激活,共同调节着细胞的生长、分化、应激、炎症反应等多种重要的生理/病理效应。
MAPK通路有4种主要的分支路线:ERK、JNK、p38/MAPK和ERK5。其中,JNK和p38功能相似,跟炎症、凋亡、生长都有关;ERK主要管细胞生长、分化,其上游信号是著名的Ras/Raf蛋白。且分支路线所使用3种激酶都是不同的,可作为通路中的Biomarker。
近二十年来,世界范围内最吸金的研究方向当属细胞信号通路与肿瘤之间的恩恩怨怨。丝裂原活化蛋白激酶MAPK信号途径是细胞中大佬级别的信息传递链,由细胞内广泛表达的Ser/Thr蛋白激酶,参与细胞的生长、发育、分化、凋亡等一系列细胞生理活动,是诱发肿瘤产生的重灾区。
目前,虽然研究MAPK信号传导与肿瘤之间关系取得了不错的进展,但要完全揭示其机制,还需要更加深入的研究。已知的MAPK信号通路有四类,今天我们主要介绍其中Ras/Raf/MEK/ERK途径与肿瘤千丝万缕的关系。
熟悉信号通路的童鞋们肯定知道Ras、Raf、MEK和ERK蛋白就是该通路中的关键因子,其中任何一个蛋白的功能异常都会导致严重的肿瘤疾病。
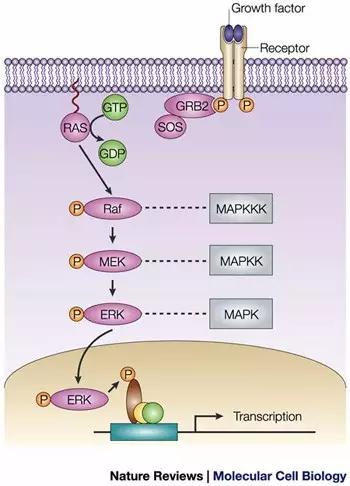
一张图记住Ras/Raf/MEK/ERK信号转导途径:
3.1、RAS蛋白与RAS*制剂抑**
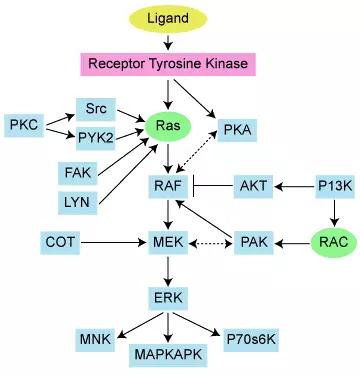
RAS蛋白就是我们平时常说的小G蛋白,当它结合GTP后具有磷酸化活性,行使下游蛋白的激活功能。RAS是多种细胞信号转导途径中的关键组分,它的活性状态对细胞的生长和分化具有重要影响。目前,在人类的癌症发生过程中,主要有KRAS、HRAS和NRAS这三类RAS蛋白。突变导致的RAS蛋白永久性激活在所有人类癌症中占据非常高的比例,故针对RAS的*制剂抑**是治疗癌症的有效药物。
RAS蛋白参与多种细胞信号转导途径:
3.2、抑癌因子FBW7与胰腺导管腺癌
原癌基因和抑癌基因时下非常热门,如YY1、FBW7、CD177和PPEF2等。接下来,我们以FBW7基因在胰腺癌中的研究成果,对“信号通路——癌基因”的研究思路有大致的了解。
FBW7基因编码的蛋白是SCF型泛素连接酶中介导底物识别的部分,常参与底物的泛素化,引导底物被蛋白酶体降解。FBW7可介导包括原癌蛋白在内的许多重要蛋白的泛素化,如cyclin E、Notch、mTOR、c-Jun和c-Myc等众多在细胞周期、细胞增殖及分化过程中发挥重要作用的蛋白质。因此FBW7的失活会引起多种恶性肿瘤的发生,如胃癌、结肠癌、乳腺癌、肝癌等。
上海肿瘤医院的虞先濬教授曾在Cell Research上发表FBW7与胰腺导管腺癌(PDAC)相关的研究性论文。其科研团队发现磷酸激酶ERK能够对FBW7进行磷酸化促进其降解,这一过程影响了FBW7发挥其肿瘤抑制因子功能,诱导胰腺癌发生。
胰腺导管腺癌(PDAC)是胰腺癌中常见的一种可怕疾病,其5年生存率不到5%,是所有癌症中致死率最高的一种。由于缺少有效的早期检测方法以及该种癌症初次诊断后的高转移率,只有15%~20%的病人能够进行切除手术。因此,提高临床治疗的实践性迫在眉睫,同时需要对PDAC发生发展的分子机制有更深入的了解。
根据早期的研究我们知道FBW7与Ras/Raf/MEK/ERK信号通路存在密切的关系,但是并不清楚FBW7的表达水平是如何受到调控的。在本研究中,科学家首先在胰腺癌临床样本中发现FBW7低表达与ERK激活显著相关,ERK激活主要是由于胰腺癌中KRAS突变导致的。
通过进一步研究发现ERK能够直接磷酸化FBW7的205位苏氨酸,促进其发生自身泛素化降解。研究人员随后证实FBW7的T205A突变体能抵抗ERK的磷酸化,并导致其中一个重要的原癌基因c-Myc表达水平下降,结果发现胰腺癌细胞增殖及肿瘤发生过程均受到显著抑制。
总的来说这些结果揭示了癌基因KRAS突变是如何通过抑制肿瘤抑制因子FBW7促进胰腺癌进展,对于胰腺癌临床实践治疗提供了良好的分子基础。
MAPK信号通路与FBW7相互调节示意图:
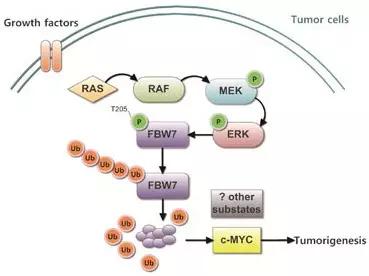
该文是如何确定FBW7的磷酸化位点及ERK对FBW7的磷酸化作用?作者根据最新的研究,确认Thr205-Pro206是FBW7上唯一的潜在磷酸化位点,然后通过两方面来确定Thr205是否为确切的激活位点:
①、FBW7蛋白上的Thr205与特异性抗体结合后,显著降低FBW7的磷酸化水平;
②、构建FBW7的T205A突变体,发现其难以被磷酸化。后续实验中,作者过表达持续磷酸化的ERK蛋白,并检测到FBW7的磷酸化水平升高而FBW7-T205A的磷酸化水平降低。
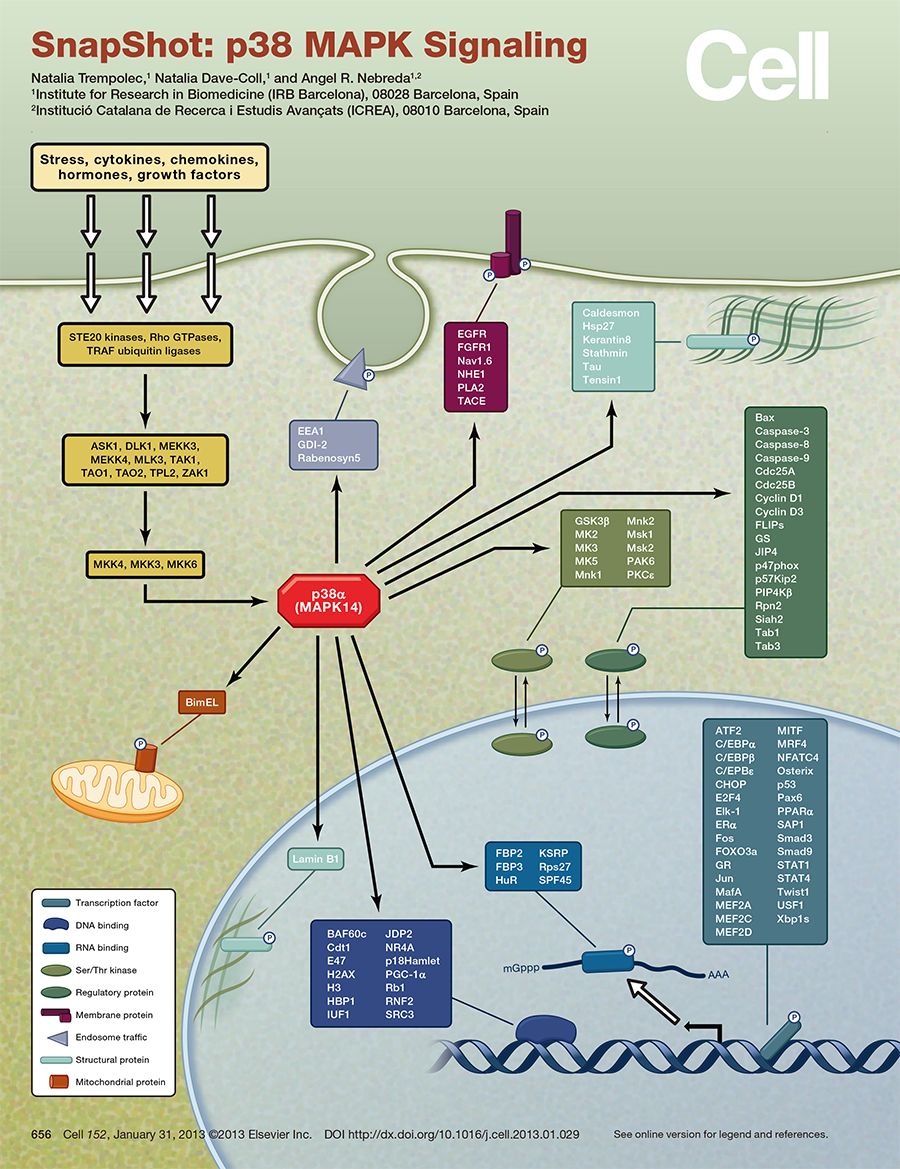
用最精炼的方式介绍MAPK 信号转导方式:胞外信号→膜受体→RAS→MAP3K→MAP2K→MAPK然后再进一步活化其他下游靶基因。上文提到,MAPK主要由四个亚家族,分别是:
①、细胞外信号调节激酶(extracellular-signalregulated protein kinase, ERK)
②、p38丝裂原活化蛋白激酶(p38 MAPK)
③、c-Jun氨基末端激酶(JNK)
④、细胞外信号调节激酶5(ERK5)
这几种MAPK亚家族参与的信号转导通路司职不同的功能,如ERK调控细胞生长和分化,JNK和p38 MAPK信号通路在炎症和细胞凋亡等应激反应中发挥重要作用。
4. JAK-STAT信号通路
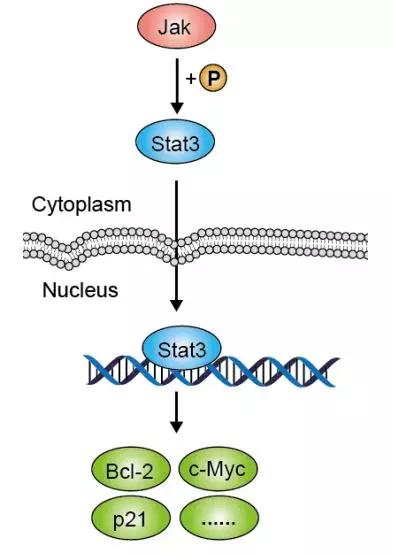
(图片截取于优秀学员Ewen的整理笔记)
JAK-STAT由三个部分组成:接收信号的酪氨酸激酶相关受体、传递信号的酪氨酸激酶JAK和产生效应的转录因子STAT。其中,磷酸化的JAK和STAT蛋白表达都是重要的Biomarker。
当多种细胞因子/生长因子与受体结合后可以磷酸化激活JAK,可磷酸化下游靶蛋白的酪氨酸残基,招募并磷酸化转录因子STAT,使其以二聚体的形式进入细胞核内与靶基因结合,调控下游基因的转录,调节细胞的增殖、分化、凋亡过程。
但值得注意的是,这些膜受体本身不具有激酶活性,但它的胞内段却存在于酪氨酸激酶JAK有结合位点,而这也是因为JAK是一类非跨膜型的酪氨酸激酶。
4.1、Jak/Stat通路的工作模式
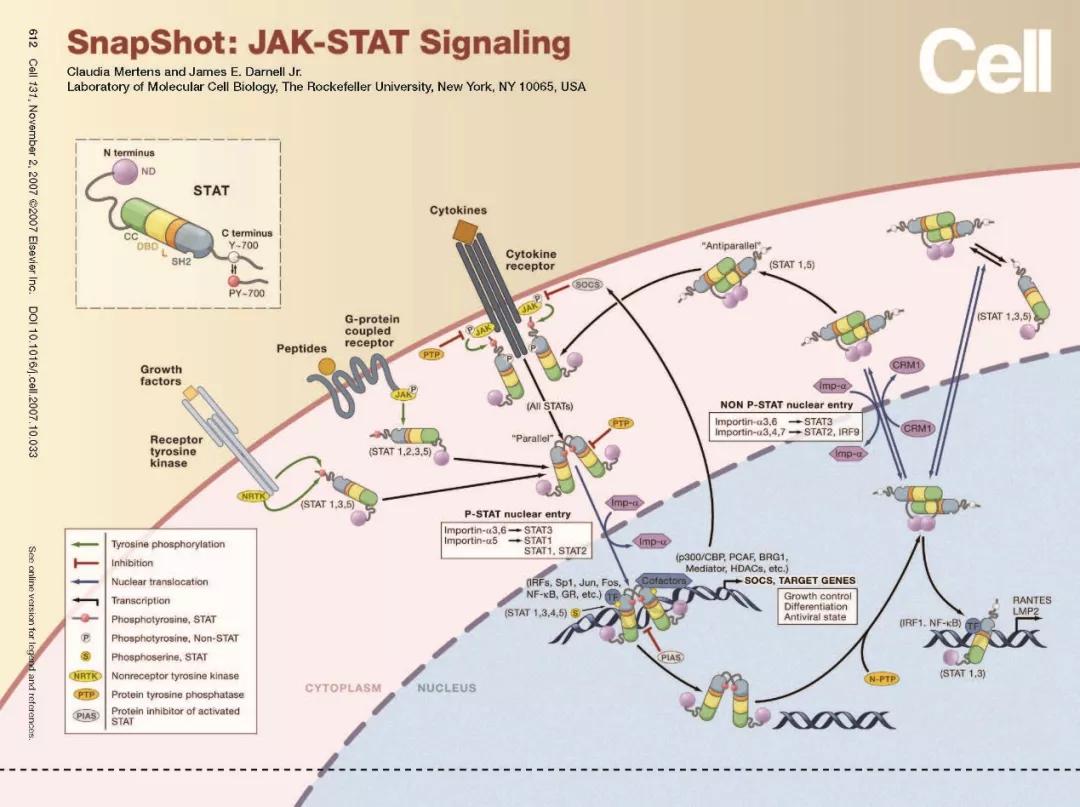
Jak/Stat通路作为应激的炎症信号通路,其反应必须要迅速。Jak和Stat是许多调节细胞生长、分化、存活和病原体抵抗信号通路中的关键部分,这一通路涉及到IL-6(gp130)受体家族,它帮助调节B细胞的分化,浆细胞生成和急性期反应。IL-6结合引起受体的二聚化同时激活受体结合的Jak蛋白,活化的Jak蛋白对受体和自身进行磷酸化。
这些磷酸化的位点成为带有SH2结构的Stat蛋白和接头蛋白的结合位置,接头蛋白将受体和MAPK、PI3K/Akt还有其他的通路联系在一起。受体结合的Stat蛋白被Jak磷酸化后形成二聚体,转移进入细胞核调节目的基因的表达。细胞因子信号传导抑制分子(SOCS)家族的成员通过同源或异源的反馈减弱受体传递的信号。
STAT在未被激活的情况下就处于细胞浆中闲逛,一旦有危险信号,马上磷酸化武装起来,迅速集结至细胞核发挥转录功能,这样的转录因子动员模式理论上说应该是非常快的,这样应对来自胞外的信号非常有利。另外,炎症中最为常见的NF-kB也是这样类型的一种转录因子,不过它是受到抑制蛋白IKBa的调控。
4.2、Jak/Stat通路的交联
Jak/Stat实际上还可以响应很多其他细胞因子的信号。JAK/STAT途径介导细胞因子的效应,除了可以用来作为抗病毒和抗增殖剂的干扰素如IL-6,还可以介导如促红细胞生成素、血小板生成素、G-CSF,这些细胞因子分别是用于治疗贫血、血小板减少症和中性粒细胞减少症的蛋白质类药物。
当细胞因子信号失调时,癌症就可能发生。此外,STAT转录因子家族还可以受到其他激酶的调节,比如mTOR、MAPK可磷酸化STAT3的Ser727使之持续激活。Stat3具有潜在促癌性(原癌基因),在许多癌症中持续的表达,这反映了细胞信号间复杂的交联。
从细胞研究的角度考虑,与其说JAK/STAT途径是炎症促进肿瘤发生的证据,不如说普遍存在的细胞内的JAK/STAT途径在长期的进化中被炎症信号选择。从逻辑上说,正常生理条件下,不被免疫反应(而非炎症反应)所动员的体细胞或其他细胞是有应对机制不被炎症因子所影响的。或许STAT家族更像一类应激因子,而非单纯的炎症通路的转录因子。
4.3、Jak/Stat通路与癌症
JAK激活突变是恶性血液病中主要的分子机制,研究人员已经在Jak2假激酶域中发现一个特有的体细胞突变(V617F),这个突变常常发生于真性红细胞增多症、原发性血小板增多症和骨髓纤维化症患者中,导致Jak2的病理激活,同时激活控制红细胞、巨核细胞和粒细胞增殖分化的促红细胞生成素(EPO)、血小板生成素(TPO)和G-CSF等的受体。而Jak1的功能获得性体细胞突变已发现存在于成人急性淋巴细胞性白血病当中。
体细胞激活突变已经证明存在于小儿急性淋巴细胞白血病(ALL)患者中。此外,在儿童唐氏综合症B-ALL以及小儿唐氏综合症患者中已发现Jak2假激酶域R683(R683G或者 ΔIREED)附近的突变。而对转录因子STAT的突变的发现却鲜有报道。有报道高免疫球蛋白E综合症(HIES)中发现STAT3基因发生的显性负效应点突变、移码突变或缺失突变。
5. TGF-β信号通路

(图片截取于优秀学员Ewen的整理笔记)
通常TGF-β信号通路的下游的效应转录因子是SMAD。该通路的信号来源有两类:TGF-β1/2/3和BMP(骨形态发生蛋白)。要检测TGF-β通路活性一般检测TGF-β、TGF-β的受体、SMAD或者是BMP即可。此外,TGF-β通路和很多通路也存在cross-talk。
TGF-β的命名是根据这种细胞因子能使正常的成纤维细胞的表型发生转化,即在表皮生长因子(EGF)同时存在的条件下,改变成纤维细胞贴壁生长特性而获得在琼脂中生长的能力,并失去生长中密度信赖的抑制作用。TGF-β信号在早期胚胎发育与组织器官形成、免疫监督、组织修复与成体稳态平衡中发挥重要作用。
TGF-β信号转导异常则可能导致多种疾病的发生,比如胚胎发育异常、肿瘤、组织纤维化、心血管疾病和免疫性疾病等。目前知道在人类*共中**存在33个TGF-β家族蛋白,包括3个TGF-β(TGF-β1/2/3)、10个BMP(Bone Morphogenetic Protein)和11个GDF(Growth and Differentiation Factor,其中GDF2=BMP9,GDF8=myostatin),以及activin、nodal、inhibin和AMH/MIS等,它们都是二聚体的分泌型多肽,根据序列、结构及信号转导分子的差异, TGF-β细胞因子大致被分为两个亚家族,其中TGF-β、activin和nodal等属于同一个亚家族,而BMP、GDF和AMH等组成BMP亚家族。
自从1981年确认TGF-β活性并于1982年成功纯化TGF-β以来,过去三十多年的研究不仅使我们发现TGF-β家族具有广泛的生物学功能,而且对于TGF-β细胞信号转导通路有了深入的认识。简单的说:TGF-β家族配体二聚体与膜上相应的II型受体和I型受体形成复合物,诱导II型受体磷酸化I型受体并激活其激酶活性,然后I型受体招募并活化下游的Smad蛋白,从而诱导Smad蛋白在细胞核内聚集并作为转录因子发挥转录调控作用。
Smad(由线虫的Sma和果蝇的Mad缩写而来)蛋白是TGF-β家族受体下游的信号转导分子。在哺乳动物中总共存在八个Smad蛋白(Smad1到Smad8)。根据在TGF-β信号转导中的功能差异,它们被分为三类:
① 包括受体调控的Smad(receptor-regulated Smad,R-Smad – Smad1/2/3/5/8)
② 通用Smad(common Smad,Co-Smad – Smad4)
③ 抑制型Smad(inhibitory Smad,I-Smad – Smad6/7)
R-Smad C末端SXS基序(X指代M或V)中的丝氨酸(S)能直接被I型受体磷酸化而导致R-Smad活化,其中Smad2/3被TGF-β/activin/nodal亚家族的I型受体ALK4/5/7磷酸化,而Smad1/5/8则主要被BMP/GDF/AMH亚家族的ALK2/3/6和ALK1磷酸化。磷酸化的R-Smad与Co-Smad/Smad4聚合并继续传递信号。抑制型Smad主要起抑制信号转导的作用(Yan and Chen, 2011)。
TGF-β家族因子
细胞信号转导过程如此简单而直接
那它们如何能够
发挥如此多样而特异的生物学功能?
首先,细胞内存在各种各样的机制对TGF-β信号转导过程进行精细调控,从而以时空特异性的方式控制TGF-β信号的强度与可持续性;
其次,为了对不同的环境因素做出不同的响应,Smad蛋白在细胞核内能与不同的转录因子或者辅助蛋白结合,从而调控不同靶基因的表达;
最后,除了经典的Smad通路,TGF-β家族细胞因子也能以细胞类型依赖的方式激活一些其他信号分子。
TGF-β细胞信号转导通路在不同的层面受到精细的调控,包括配体、受体、Smad以及核内转录水平的调控;其调控机制多种多样,比如蛋白-蛋白相互作用、蛋白翻译后修饰、蛋白降解、蛋白运输与细胞内定位,以及Smad-DNA结合等。
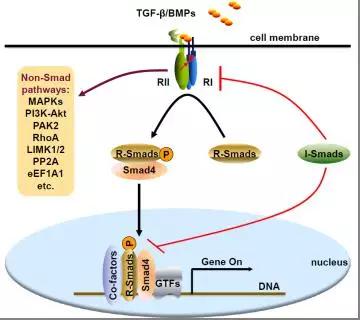
TGF-β家族细胞因子诱导细胞膜上的丝氨酸/苏氨酸激酶型受体形成功能性复合物:两个II型受体(RII)和两个I型受体(RI)。
RII磷酸化RI胞内结构域中的GS区
▼
激活RI的激酶活性
▼
RI继续磷酸化下游R-Smad蛋白C末端的SXS基序
▼
诱导Smad复合物形成、细胞核转运以及Smad-DNA结合
▼
Smad与通用转录因子(general transcriptional factor,GTF)、其它转录因子或者辅助蛋白一起调控靶基因转录。
TGF-β家族细胞因子也能以细胞类型依赖的方式激活其它信号分子:MAPK、PI3K-Akt、PAK2和RhoA等。除了R-Smad和Co-Smad/Smad4,抑制型Smad蛋白(I-Smad,包括Smad6和Smad7)是TGF-β信号通路的关键负调控因子,它们既能在细胞质中抑制受体/Smad活性,也能作为转录抑制蛋白在细胞核中发挥作用。
6. Wnt信号通路

(图片截取于优秀学员Ewen的整理笔记)
TGF-β和Wnt都是调节上皮间质转化EMT的至关重要通路。一般Wnt通路主要指的是由β-Catenin介导的经典信号通路。β-Catenin是检测Wnt是否激活的一个重要Biomarker。
而在跨膜受体FZD蛋白家族接收Wnt信号后,可通过下游蛋白激酶的磷酸化作用抑制β-Catenin的降解活性,随后胞浆中稳定积累的β-Catenin进入细胞核后结合TCF/LEF转录因子家族,启动下游靶基因的转录。
6.1、Wnt信号通路的构成
Wnt信号通路是一个复杂的调控网络,目前认为它包括三个分支:
经典Wnt信号通路,即Wnt/β-cantenin信号通路
Wnt/PCP通路 (planner cell polarity pathway)
Wnt/Ca2+通路,由Wnt5a和Wnt11激活
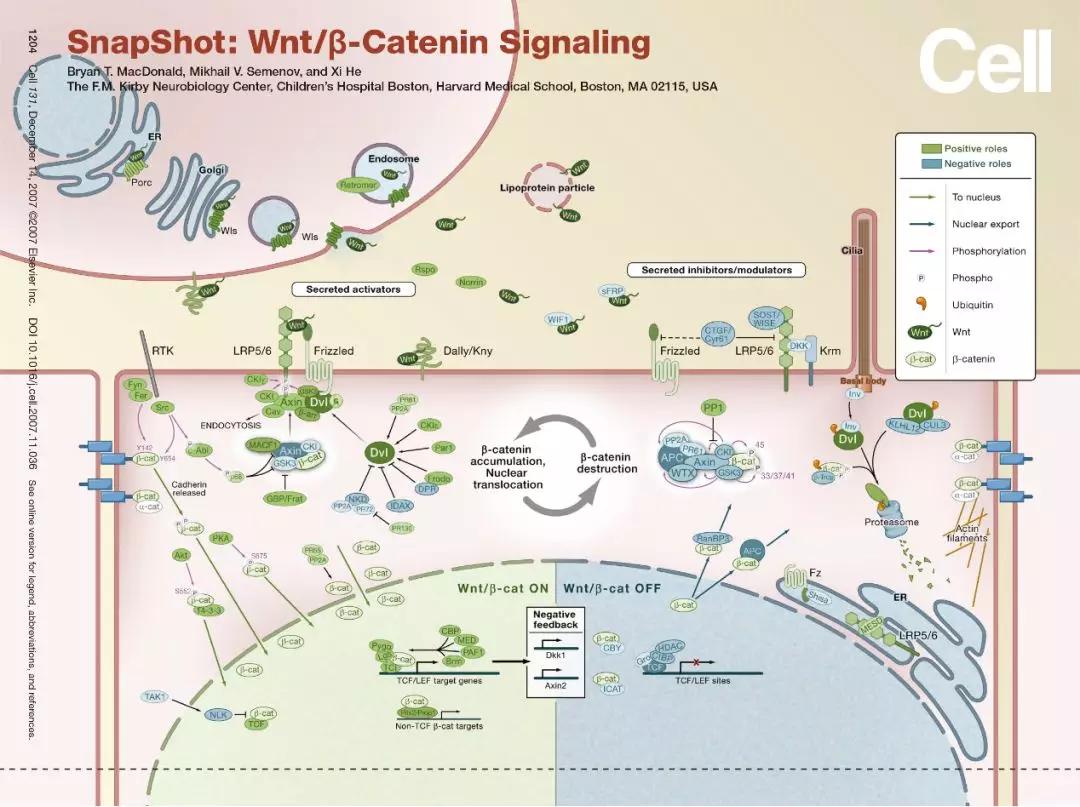
这3条信号通路主要成分包括:分泌蛋白Wnt家族、跨膜受体Frizzled家族、CK1、Deshevelled、GSK3、APC、Axin、β-Catenin、以及转录因子TCF/LEF家族。不过我们一般提到Wnt信号通路,主要指的还是由β-catenin介导的经典Wnt 信号通路,下面我们就此简单介绍下Wnt信号通路的主要组成部分:
①、Frizzled (Fzd或Frz)
分泌型糖蛋白Wnt的细胞膜上受体,为7次跨膜蛋白。FZD胞外的N端有一个富含半胱氨酸的结构域(cysteine rich domain, CRD),能与Wnt结合。
②、Dishevelled (Dvl)
Dvl蛋白在细胞质中接受上游信号,通过抑制APC、Axin以及GSK3β等蛋白形成的复合物的功能,稳定细胞质中游离状态的β-Catenin蛋白。细胞质中积累的β-Catenin蛋白进入细胞核与TCF/LEF家族的转录因子结合,从而开启了下游靶基因的转录。
③、GSK3β
是一种丝氨酸/苏氨酸蛋白激酶。在没有Wnt信号时,GSK3β能将磷酸基团加到β-Catenin N端的丝氨酸/苏氨酸残基上,磷酸化的β-Catenin经β-TRCP泛素化共价修饰后,被蛋白酶体(proteasome)降解。
④、酪蛋白激酶1 (casein kinase 1,CK1)
能将β-Catenin的Ser45位点磷酸化,随后GSK3β将β-Catenin的Thr41、Ser37、Ser33位点磷酸化。
⑤、Axin
是一种支架蛋白,具有多个与其它蛋白作用的位点,能与APC、GSK3β、CK1等形成β-Catenin降解复合物。此外它还与Dvl、PP2A等Wnt信号的其它组分相互作用。
⑥、TCF/ LEF
是一类具有双向调节功能的转录因子,它与Groucho结合可以抑制基因转录,而与β-Catenin结合则促进下游靶基因的转录。
简单来说,当细胞没有接受Wnt信号刺激时 (Wnt Off),细胞质内的Axin、APC和GSK3β形成毁灭复合体(destruction complex),与β-Catenin结合并使其被磷酸化后,最终通过泛素化修饰而降解。
在Wnt激活过程中,Dvl是一个重要的效应分子,当Wnt与其膜受体FZD结合后 (Wnt On),激活胞内蛋白Dvl并抑制GSK3β等蛋白形成的β-Catenin降解复合物的降解活性,稳定细胞质中游离状态的β-Catenin蛋白。胞浆中稳定积累的β-Catenin进入细胞核后结合LEF/TCF转录因子家族,启动下游靶基因 (如c-myc、Cyclin D1等) 的转录。
6.2、Wnt通路的交联
Wnt/β-catenin 途径调控着干细胞的多能分化、器官的发育和再生,在功能上与Hippo、Notch和TGF-β等类似,当然也与这些发育调控相关的信号通路有着不同程度的交联。在这里,主要简单介绍下Wnt与Notch之间的“基情”。
在小肠干细胞和小肠稳态调节过程中,Notch和Wnt信号扮演了重要的调控作用,阻断Notch信号会引起小肠干细胞分化失衡,而在这一过程中同时解除了对Wnt通路的抑制。器官的稳态平衡需要对成体干细胞严格调控,并通过多种信号整合实现干细胞向终末细胞的分化。
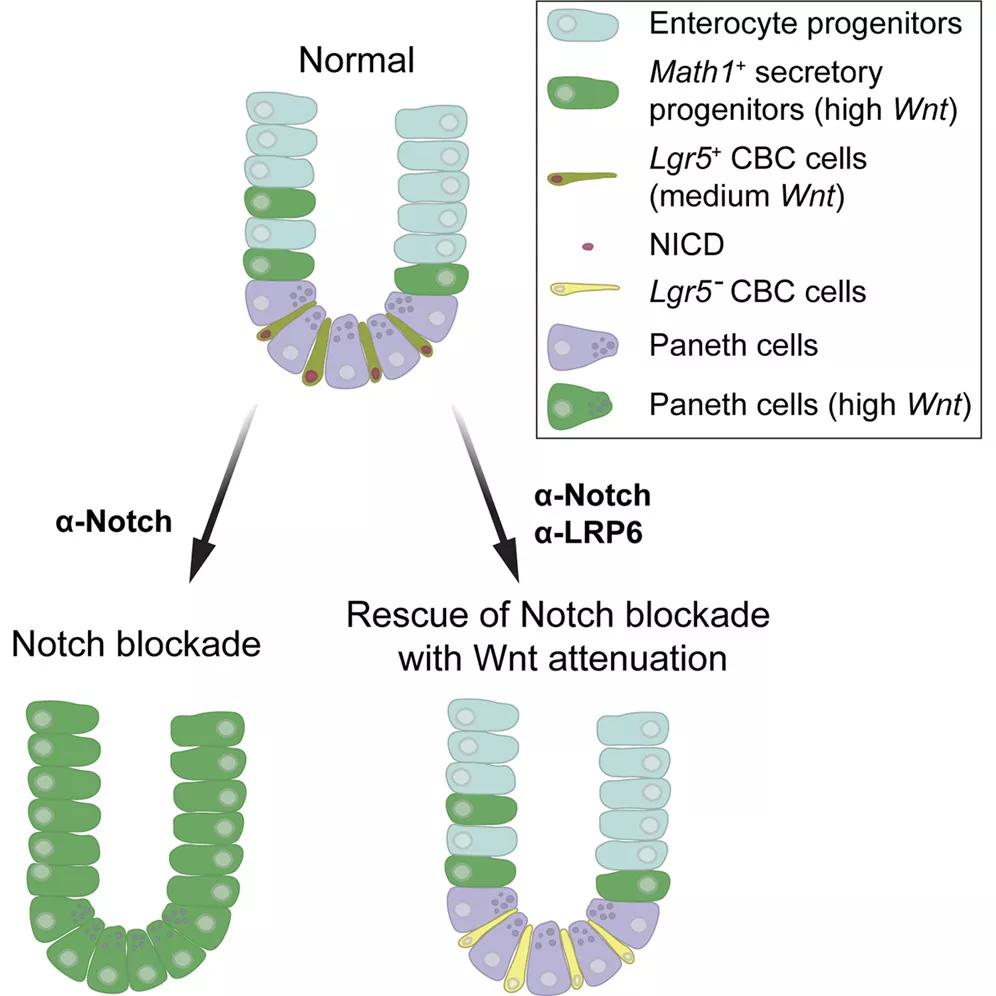
Tian, H., et al., Opposing activities of Notch and Wnt signaling regulate intestinal stem cells and gut homeostasis. Cell Rep, 2015. 11(1): p. 33-42.
当Notch信号受到抑制,小肠干细胞倾向于分化成分泌细胞,并抑制营养吸收细胞的分化趋势。当Notch受体被特异性抗体阻断后,结果发现阻断Notch会解除Wnt途径的抑制作用,扰乱小肠干细胞的正常功能,导致促分泌功能相关基因的错误表达。Wnt与Notch信号之间的相互作用,对维持干细胞活性及分化平衡的负向调节有着重要的生理意义。
综上所述,人们自然而然地选择把Wnt/β-Catenin信号通路中的关键蛋白作为药物靶点,筛选分子药物治疗癌症。目前已经有多种Wnt通路靶向*药性**物进入临床阶段。不过,正如我们一直在强调的,癌症的发生是一个多因素、多阶段、多基因变异积累的复杂过程,多种信号通路可能同时参与了癌症的发生。随着未来研究的深入,期待更多Wnt信号通路新成员的发现。细胞内信号通路相互协同机制的研究,能够为我们设计更加有效的抗癌药物提供更多的理论基础。
7. Notch信号通路
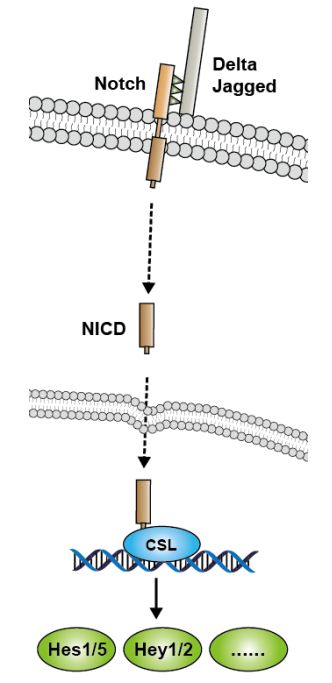
(图片截取于优秀学员Ewen的整理笔记)
Notch跟以上通路的两个显著区别:
①、Notch的受体和配体都是膜蛋白,它介导的是两个细胞相互靠近接触之后的活化效应,而不是由分泌型的蛋白作为配体。
②、Notch通路不是通过激酶磷酸化逐步活化传递信号,它是Notch通过三步蛋白酶切水解,把有转录调节活性的Notch蛋白片段(NICD或ICN)释放出来,再与转录因子CSL结合,调节下游基因表达。
7.1、恶性肿瘤的本质是细胞的不正常增生,局部侵入周围正常组织,甚至经循环系统转移到身体的其他部位。从分子病理学的角度来说,恶性肿瘤的根源是DNA突变的积累。而突变的积累则导致促进细胞生长的蛋白质大量表达,使得细胞周期控制失常。其中,Notch信号传导通路中的某些分子的DNA突变与恶性肿瘤的发生发展有着广泛而密切的关系。
Notch信号传导通路是决定细胞命运的最重要的通路之一。Notch 通路主要由四部分组成:Notch受体、Notch配体、CSLDNA结合蛋白、下游靶基因。上述四部分中任何一个因素改变都会对Notch信号通路传递产生影响。
相邻细胞间通过Notch受体传递信号,可以调节细胞的分化,增殖和凋亡。Notch受体是由Notch基因编码的单跨膜蛋白,在包括人类在内的哺乳动物中发现了4种Notch受体(Notch 1,2,3,4)。
而Notch配体也是表达于细胞表面的单跨膜蛋白,相邻细胞通过Notch受体和配体的结合,传递Notch信号。人体有5种Notch配体,分别是Jagged 1、Jagged 2、Delta 1、Delta 3和Delta 4。
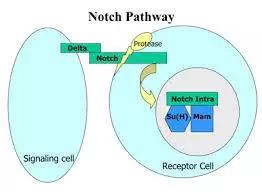
Notch配体与受体结合后触发Notch信号的活化,Notch受体相继发生2次蛋白水解,并进一步转移到细胞核,从而激活靶基因的转录,发挥生物学功能。Notch信号调节机体正常发育的一些关键步骤,所以,该通路的某些分子发生突变,或者其下游事件发生改变,都会导致恶性肿瘤的发生。
7.2、Notch信号传导通路与恶性肿瘤的关系首先在人类急性T淋巴细胞白血病(T-ALL)中得到了证实。研究发现,几乎所有的T-ALL都高表达Notch 1或者Notch 3。而最新的研究发现,大部分T-ALL患者体内都存在Notch 1的活性突变。

另外,Notch受体及配体在多种恶性肿瘤细胞中均有大量表达。在非小细胞肺癌(NSCLC)中,Notch 1和Notch 2存在高度表达的情况。除了对NSCLC细胞的增殖具有高度的促进作用,Notch信号传导通路还可以介导NSCLC通过循环系统和淋巴系统转移到人体的其他部位。
在神经胶质细胞瘤中,发现Notch 1以及Notch配体Delta 1和Jagged 1高度表达。在肾脏细胞癌中,发现Notch 3大量表达。
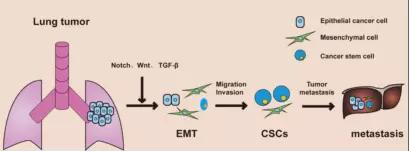
致癌事件不一定仅仅发生在Notch基因上,也可以发生在Notch信号传导通路的下游。研究发现,EB病毒可以产生一种名为EB核抗原2(EBNA2)的蛋白质,该蛋白质可以模拟胞内域(intracellular Notch,ICN),进入细胞核,在相关辅助因子的参与下,与CSL蛋白相结合并使之活化,激活下游的靶基因的转录,诱发恶性肿瘤。
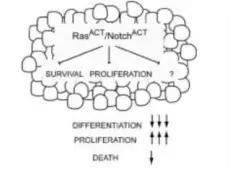
7.3、在胰腺癌的病人中,发现了Notch信号传导系统的活化,和Notch受体和配体的大幅度上调。另外,Notch也可以作为致癌基因Ras的帮凶促进上皮细胞癌的发生发展。
如下图所示,在Notch和Ras的共同作用下,细胞的分化程度大大降低,而增殖程度大大增加,其最终结局就是恶性癌变。
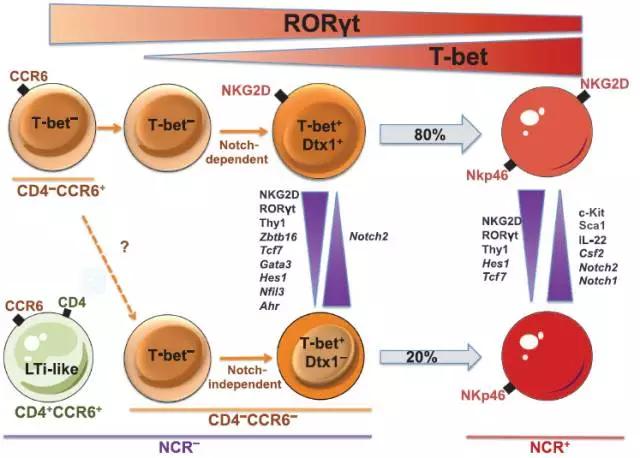
Science中的一篇论文提到小鼠体内固有淋巴细胞由两个亚群组成:NCR+ ILC3s和NCR- ILC3s。在小肠固有层中,有80%的NCR- ILC3s是依赖于Notch2的活化来刺激转录因子T-bet,RORγt和芳烃受体AhR基因的表达,进而转化为NCR+ ILC3s细胞。
然而,由于NCR- ILC3s细胞转变为NCR+ ILC3s细胞部分是由AhR和T-bet的表达量控制的,因而仍有20%NCR+ ILC3s的生长发育是由NCR- ILC3s通过T-bet依赖途径转化而来的。这部分细胞亚群的生长及转录因子的丰度增加均无需依赖Notch信号,但是其细胞因子的分泌及表面NCR的表达却依赖于Notch信号。由此文章得到的推论:Notch信号通过控制NCR+细胞命运来调节ILC3s的可塑性。
NCR- ILC3细胞在通过Notch依赖的信号途径转变为NCR+ ILC3时,其胞内的Ncr1曾进行了短暂表达,即具有NCR- ILC3(FM+)表型。然而,在肠道中,由Treg细胞分泌的转化生长因子(TGF-β)通过与其受体(TGF-βRI&TGF-βRII)结合后,可以抑制NCR- ILC3s细胞向NCR+ ILC3s细胞转化。因而,Notch和TGF-β可相互调节NCR- ILC3s和NCR+ ILC3s亚群之间的平衡,以确保肠道内环境的稳定。
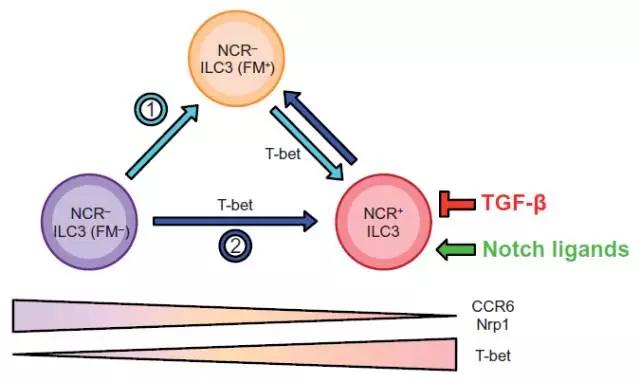
恶性肿瘤的发病机制非常复杂,到目前为止,我们还不能说已经完全了解和掌握了它。但是,可以明确的是Notch信号传导通路广泛参与了恶性肿瘤的发生发展。Notch信号传导通路发挥致癌作用,导致细胞在增殖,细胞周期抑制,分化,和凋亡的过程失调和失控,造成了细胞的转化及恶性化,最终导致了恶性肿瘤的发生。
8. Hedgehog信号通路
(图片截取于优秀学员Ewen的整理笔记)
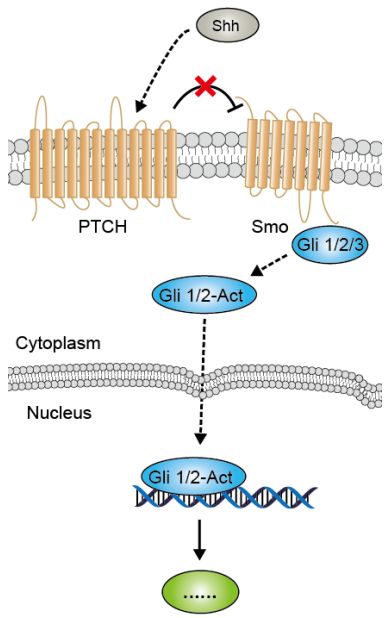
Hedgehog通路的配体就是Hh蛋白,需要经过一系列加工才有活性。Hedgehog通路的受体有两种,一种是Ptc,一种叫Smo。该通路的特点是内部具有反馈调节机制——正常情况下Smo是正向激活转录因子,但另一个受体Ptc能抑制Smo蛋白活性,可抑制下游通路,实现自我调节。Hedgehog通路的转录因子是Gli蛋白家族,有几种不同亚型,Gli1是常见的检测对象。另,这个通路与Wnt、Notch通路的cross-talk也是非常密切的。
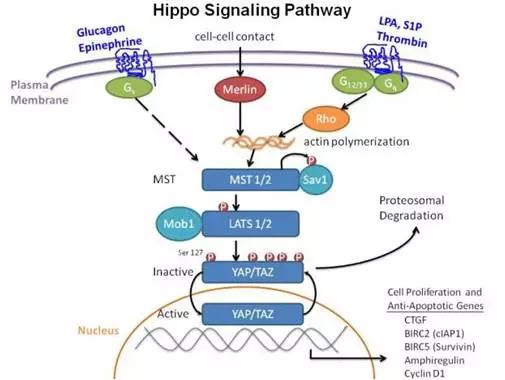
9. Hippo信号通路
(图片截取于优秀学员Ewen的整理笔记)
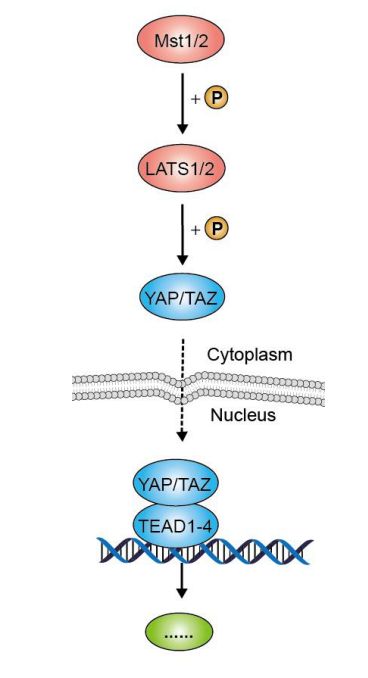
Hippo信号通路由一组保守的激酶构成,是一条抑制细胞生长的信号通路。在哺乳动物中,Hippo信号通路上游的膜蛋白受体感受到胞外环境的生长抑制信号后,经过一系列激酶的磷酸化反应,最终作用于下游效应因子YAP和TAZ。YAP和TAZ继而与细胞骨架蛋白相互作用,被滞留在胞质内,不能进入细胞核行使其转录激活功能,从而实现对器官大小和体积的调控。
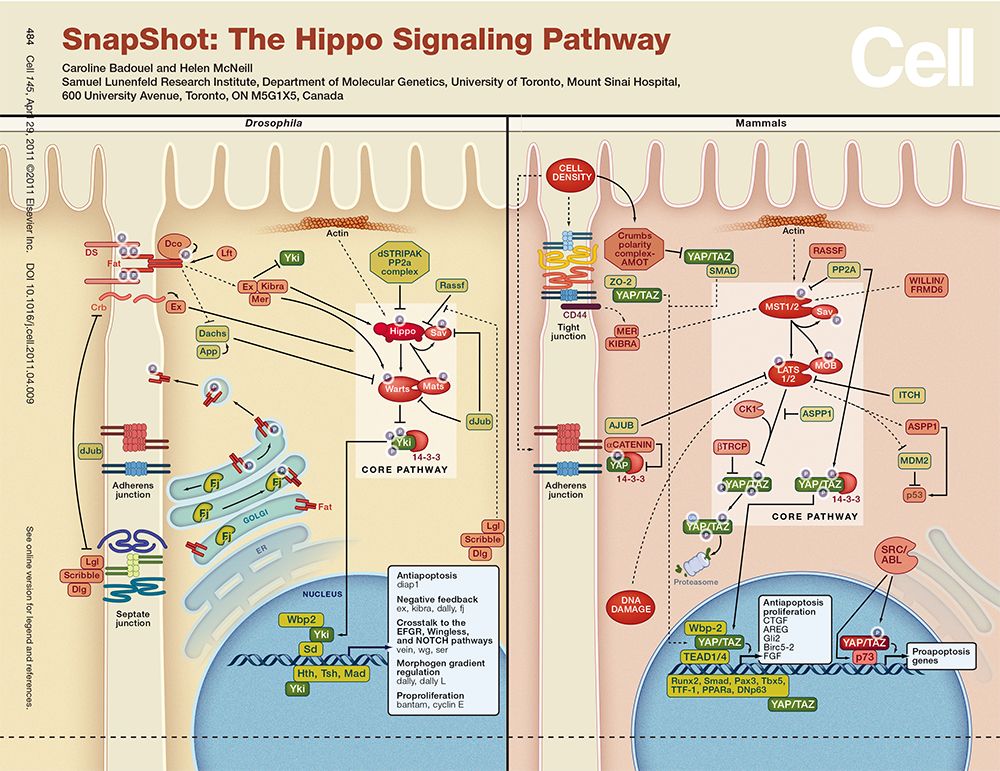
近年来的研究证实:Hippo信号传导通路还在癌症发生、组织再生以及干细胞的功能调控上发挥着重要的作用。下面,我们就来详细的看看Hippo信号通路的作用机制。
9.1、YAP和TAZ对骨细胞分化的调控
Runt相关转录因子2(RUNX2)是一种刺激成骨作用的基本转录因子。研究发现, TAZ与RUNX2结合能强有力地促进骨发生程序的执行。将鼠骨髓来源MSCs中的TAZ敲低后在成骨条件培养, 会出现钙沉积作用丧失, 成骨细胞分化缺陷。在C2C12细胞中敲除TAZ可抑制其分化为成骨细胞, 而TAZ水平增加又会促进其向骨细胞分化。
YAP在MSCs向骨细胞分化中的作用还未完全阐明。YAP含有一个SH3结合结构域, 可以直接与非受体酪氨酸激酶Src结合, 而TAZ缺少这一SH3结合结构域。有研究认为, YAP也可作为RUNX2的共激活因子, 核定位的YAP突变体能促进骨形成。
然而, 还有研究发现, YAP在应答于Src刺激时, 可作为骨钙素启动子上RUNX2的共抑制因子发挥功能。YAP与RUNX2的结合受到Src介导的YAP酪氨酸磷酸化促进, 在人骨髓来源的MSCs中, 抑制Src活性会刺激成骨细胞分化。因而, Hippo信号通路与酪氨酸激酶信号通路交叉对话在骨细胞分化中的作用还需进一步研究。
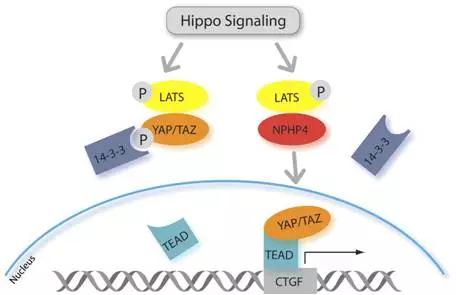
9.2、TAZ对MSCs向脂肪细胞分化的调控
MSCs的另一个命运决定方向为向脂肪细胞分化, Hippo信号通路在这一过程发挥核心作用。在3T3-L1脂肪祖细胞或鼠骨髓来源MSCs中, 敲低TAZ并在促脂肪生成条件培养, 脂肪细胞形成增加, 表明TAZ作为脂肪形成的负向调控因子发挥功能。过氧化物增殖子激活型受体γ(PPARγ)是核激素受体超家族成员, 在脂肪形成过程中作为一个关键转录因子发挥功能。
TAZ通过结合到PPARγ的PPXY模序上抑制PPARγ活性。在脂肪形成过程中YAP是否以相似方式发挥功能目前还未进行研究。有趣的是, MST1和MST2激酶可促进SAV1与PPARγ结合, 这种结合是由SAV1的WW结构域与PPARγ的PPXY模体介导。
在3T3-L1细胞中, SAV1表达会稳定PPARγ蛋白水平, 而SAV1敲低会导致PPARγ水平降低。SAV1与PPARγ结合可能阻断了PPARγ被泛素化通路元件接近, 并阻止蛋白酶体对PPARγ的降解。
事实上, SAV1可能也与TAZ竞争结合PPARγ, 因为这两个蛋白结合在同一个PPXY模体上。由于MST1/2与SAV1刺激LATS1/2的活性, 它们还可能通过促进TAZ磷酸化使其定位于细胞质, 间接影响TAZ与PPARγ在细胞核中结合。
SAV1是否通过调控TAZ发挥功能目前还不清楚, 但它在脂肪形成过程中的作用表明Hippo信号通路其他元件也参与了这一过程。

9.3、YAP和TAZ和对MSCs向肌细胞分化的调控
成体骨骼肌的再生主要通过肌卫星细胞增殖与分化完成, 肌发生分化因子1(Myo D)、肌肉调节因子4(Mrf 4)以及myogenin等转录因子在肌卫星细胞分化中起作用。研究发现, 除肌卫星细胞外, 存在于成体许多部位中的MSCs也能支持骨骼肌再生。
在肌卫星细胞中, 异位过表达TAZ会以一种Myo D依赖性方式增强生肌基因表达并加速肌纤维的形成, 而TAZ敲低会延迟生肌分化。与此类似, 在MSC样细胞中, TAZ与Myo D共表达会加快Myo D诱导的生肌性分化。
进一步研究显示, 在MSC样细胞和成肌细胞中, TAZ通过WW结构域与Myo D相互作用, 增强Myo D与myogenin基因启动子结合, 激活myogenin和MCK基因转录, 诱导肌细胞终末分化的发生。
综上所述,Hippo信号通路是一条抑制细胞生长的信号通路。近年来的研究证实:Hippo信号传导通路还在癌症发生和转移、组织再生以及干细胞的功能调控上发挥着重要的作用。Hippo信号传导通路的研究正越来越受到分子生物学领域的关注。
虽然通路这个变量包含大量的标,但看惯了这些通路里面的明星分子,就熟悉了基础科研的一些套路。
>>>>>>>>>>>>>>>>>>>>>>>>>>>>>>>>>>>完<<<<<<<<<<<<<<<<<<<<<<<<<<<<<<<<<<<<<<
10天领悟3分SCI套路训练营
点击下方了解详情报名