原创 张腾倩等 南京林业大学学报

论文推荐
欧美杨 PdMODD 基因克隆与表达特性分析
张腾倩 1 ,张伟溪 1 ,丁昌俊 1 ,张 静 1 ,,胡赞民 2 ,苏晓华 1 *
1. 林木遗传育种国家重点实验室,国家林业和草原局林木培育重点实验室,中国林业科学研究院林业研究所;2.中国科学院遗传与发育生物学研究所。

近年来,随着RNAi、基因编辑技术的不断发展,转录抑制因子基因挖掘已成为基因工程育种研究的热点之一。迄今为止,在植物中鉴定的最具代表性的转录抑制基序为EAR ( ethylene-responsive element binding factor-associated amphiphilic repression)基序,其包含一段保守序列LxLxL或DLNxxP,广泛存在于各种转录抑制因子中1EAR基序介导的转录抑制被认为是与转录激活并存且极为重要的一种基因调节机制,研究表明,含有EAR基序的蛋白可以通过负调控基因表达在植物器官发育.激素应答、抗病反应及干旱胁迫响应等生物学过程发挥重要的作用。
MODD( mediator of OsbZIP46 deactivation and degradation)是由Tang 等首次在水稻中鉴定的具有典型EAR基序的转录抑制因子,属于NINJA 家族蛋白。NINJA家族蛋白序列一致性高,都含有A、B、C 3个保守区域,其中A保守区域为EAR基序,该基序是NINJA家族蛋白与协同抑制因子TPL( TOPLESS)和TPRs ( TPL-related proteins)的互作位点;B区域含有一段核定位信号序列;C区域为NINJA蛋白与JAZ( jasmonate zim-domain)互作的位点。
研究表明,水稻MODD基因在ABA信号传导和干旱胁迫应答中具有负调控作用:干旱胁迫下,MODD过表达株系的存活率显著低于野生型,而突变株系增加了对ABA的敏感性和抗旱性;MODD可通过EAR基序结合OsTPR3蛋白招募表观调控因子HDA702抑制OsbZIP46(ABA信号传导正调控因子)活性,通过OsPUB70介导的泛素化抑制OsbZIP46稳定性,进而调节水稻ABA信号传导和抗旱性。此外,MODD还能与水稻中其他干旱正调控因子OsbZIP23和OsbZIP7210相互作用。但是目前关于MODD基因在木本植物中的研究还鲜见报道。
‘渤丰3号’杨,雌株,属欧美杨,是中国林业科学研究院林业研究所自主培育的短轮伐期速生优良品种,已获得国家林木良种审定,具有速生、适应性强等特性,适于辽宁、山东、河南等地生长。我国干旱、半干旱地区面积广阔,生态环境恶劣,盐碱地总面积达9900万hm²,因此挖掘林木抗逆胁迫相关基因具有重要意义。 本期论文推荐的作者 研究基于水稻 MODD 氨基酸序列,通过同源比对的方法在欧美杨中得到3条 PdMODD 基因,并对其序列特征、亲缘进化关系、组织特异性及NaCl和聚乙二醇(PEG)胁迫下的表达模式进行分析,为进一步探究杨树 MODD 基因功能提供理论依据,并为培育优良抗逆杨树新品种提供候选基因资源。
下面跟学报君一探究竟!
作者简介
通讯作者

苏晓华,本刊编委 。中国林业科学研究院林业研究所研究员(二级),林木育种首席专家,主要从事林木遗传育种研究。
第一作者

张腾倩,女,1993年4月出生,林木遗传育种国家重点实验室林木遗传育种专业博士研究生。

关键词 : ‘渤丰 3号’杨; PdMODD ;生物信息学分析;表达模式分析
基金项目: 国家转基因生物新品种培育科技重大专项课题(2018ZX08020002)。
引文格式: 张腾倩,张伟溪,丁昌俊,等.欧美杨PHMODD基因克隆与表达特性分析[J].南京林业大学学报(自然科学版),2021,45(2):43-50.ZHANGTQ,ZHANGWX,DINGCJ,et al. Cloning and expression characlteristics of PdMODD genes in Populus × euramericana [J]. Journal of Nanjing Foresty University (Natural Sciences Edition) ,2021,45(2):43-50.DOI:10.12302/j.issn. 1000-2006.202007015.

1目的
研究杨树重要胁迫响应新基因,为揭示杨树抗逆分子机制、培育优良抗逆杨树新品种提供理论依据。
2方法
基于水稻MODD氨基酸序列,通过BLAST在美洲黑杨基因组中筛选得到3条基因(Podel.08G114100,Podel.10G158900和Podel.17C098500),并以其序列为参考,以‘渤丰3号’杨cDNA为模板克隆得到3条基因( PdMODD 1 、PMODD 2和 PdMODD 3)。利用生物信息学分析 PdMODD 蛋白的结构特征及与其同源蛋白的亲缘关系。通过基因枪轰击法分析 PdMODD 基因亚细胞定位。采用实时荧光定量(qRT-PCR)技术,分析 PdMODD 基因组织特异性和不同胁迫条件下的的表达模式。
2.1 试验材料
实验材料为中国林科院林业研究所杨树课题组自主选育的优良新品种‘渤丰3号’杨。将生长40d的组培苗炼苗后置于光照时间为16 h、平均温度25℃、相对湿度70%~80%的温室内进行水培,培养液为1/10 Hoagland营养液,培养1周后选取生长状态良好且长势一致的‘渤丰3号’杨,分别用含有200 mmol/L NaCl和质量分数20%聚乙二醇(PEGoo)的培养液进行处理,同时设以正常培养液培养的对照,在处理0、3、6、12、24、48 h时进行根、茎、叶组织的取材,材料液氮速冻后于-80 °C冰箱保存备用。每个处理,至少3株,3次重复。
2.2 研究方法
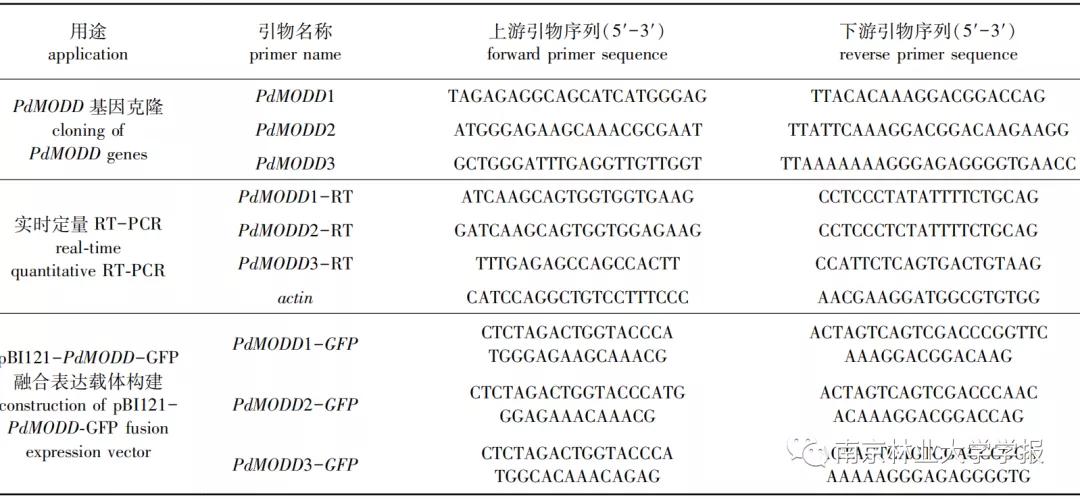
基于水稻MODD氨基酸序列,以美洲黑杨( Populus deltoides WV94 v2.1)为参考基因组,利用JGl( https :// phytozome.jgi.doe.gov/ pz/ portal.html#!search? show =BLAST)进行BLAST,筛选出同源性 较高(期望值<1×10-10)的3条基因Podel.08G114100、Podel.10G158900和Podel.17G098500,分别以3条基因的CDS序列为参考设计引物,引物序列见 表1 。以‘渤丰3号’杨cDNA为模板进行PCR扩增,得到的基因序列分别命名为 PdMODD 1、 PdMODD 2和 PdMODD 3 (GenBank登录号:MW255958、MW255959、MW255960)。其中PCR反应体系为:PCR Mix 10 μL,上下游引物(10μmol/L)各1 μL,模板2 μL,去离子水6 μL;反应程序为:95 ℃ 3 min;95 ℃ 30 s,58 ℃ 30 s,72 ℃l min,35个循环;72 ℃ 10 min。扩增产物经琼脂糖凝胶电泳检测,回收纯化后连接pMD18-T载体,转化大肠杆菌感受态DH5a,挑取阳性单克隆经菌液PCR验证后进行测序。
利用NCBI中CD-search ( https :// www.ncbi.nlm.nih.gov/Structure/ cdd/wrpsb. cgi)预测PdMODD蛋白序列的保守结构域。用ProtParam( http:// au.expasy.org/tools/ protparam.html)计算推导 PdMODD 基因所编码蛋白质的分子质量、理化等电点不稳定系数和亲水性平均系数。用Plant-mPIoc ( http://www.csbio.sjtu. edu.cn/ bioinf/ plant-multi/)预测PdMODD蛋白的亚细胞定位。利用BioEdit对PdMODD的同源蛋白序列进行多序列比对;通过MEGA5.05软件使用Neighbor-Joining(NJ)法对PdMODD及其同源蛋白进行系统进化分析,构建系统进化树。
▼表 1 研究设计的引物序列
3结果
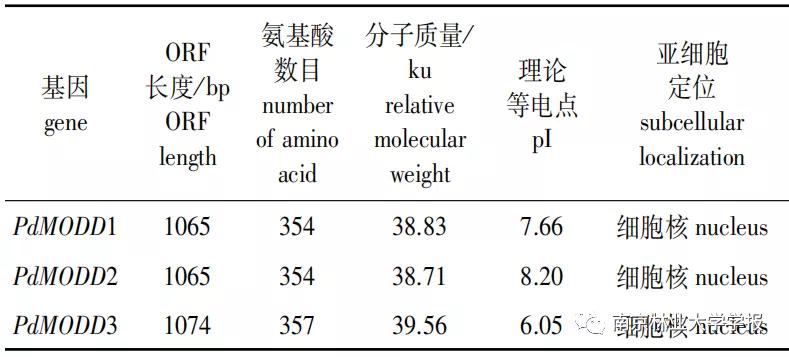
PdMODD 1—3基因的开放阅读框(ORF)全长分别为1065,1 065和1074 bp,编码354,354和357个氨基酸,均为不稳定的亲水性蛋白。 PdMODD 基因均系统进化树分析显示, PdMODD 1蛋白与毛果杨PAFP2亲缘性较高并与拟南芥AtAFPl和木薯MeAFP2位于同一个分支; PdMODD 2蛋白与麻疯树JcAFP2亲缘关系最近,并与拟南芥AtAFP2聚成一个分支; PdMODD 3与毛果杨PtAFP3-1亲缘关系最近,与栓为NINJA家族成员,含有3个保守结构域,其中一个为转录抑制基序EAR。系统进化树分析显示, PdMODD 1蛋白与毛果杨PtAFP2亲缘性较高并与拟南芥AtAFPI和木薯MeAFP2位于同一个分支; PdMODD 2蛋白与麻疯树JcAFP2亲缘关系最近,并与拟南芥AtAFP2聚成一个分支; PdMODD 3与毛果杨PtAFP3-1亲缘关系最近,与栓皮槠QsAFP3、拟南芥AtAFP4属于同一分支。亚细胞定位结果表明 PdMODD 基因均定位于细胞核。组织特异性分析显示, PdMODD 1—3在根、茎、叶中均能表达,且在叶中表达量最高,根中表达量最低。胁迫响应表达模式结果显示,NaCl和聚乙二醇(PEG)胁迫处理下, PdMODD 1—3在根、茎、叶组织中主要为显著上调表达。
3 .1 PdMODD 基因的克隆与序列分析
以‘渤丰3号’杨cDNA为模板,以在美洲黑杨基因组中筛选的3条基因(Podel.08G114100、Podel.10G158900和Podel.17G098500)CDS序列为参考设计引物,PCR扩增得到3条基因,分别命名为 PdMODD 1,扩增产物于琼脂糖凝胶电泳检测( 图1) 。胶回收产物纯化后连接pMD18-T,转化大肠杆菌感受态DH5a,将菌液PCR检测得到的阳性克隆进行测序,测序获得的 PdMODD 1—3基因序列与美洲黑杨基因组中对应的序列相似度很高,分别为97%、99%和99%。
对 PdMODD 基因序列进行分析,结果显示 PdMODD 1—3的开放阅读框(ORF)长度分别为1065、1 065和1 074 bp,分别编码354、354和357个氨基酸 (表2) 。理化性质分析显示PdMODD分子质量分别为38.83、38.71和39.56 kU ,理论等电点为7.66,8.20和6.05,不稳定系数为60.31,64.18和64.12,亲水性平均系数为-0.847、-0.873和-0.775,均为不稳定的亲水性蛋白。亚细胞定位预测推断3条 PdMODD 基因编码产物均定位于细胞核。

▲图 1 ‘渤丰 3号’杨 PdMODD 基因的克隆
▼表 2 PdMODD 基因特性分析
3.2 PdMODD蛋白保守结构域及进化树分析
利用NCBI 中CD-search分析PdMODD蛋白保守结构域,结果表明PdMODD蛋白均含有EAR、NINJA_B和Jas保守结构域,属于NINJA家族。通过NCBI网站进行BLAST比对,发现PdMODD与NINJA家族AFP蛋白同源性较高,选择相似度较高的毛果杨、麻疯树、木薯等6种植物AFP氨基酸序列进行比对,结果显示,PdMODD与其他6种植物AFP氨基酸序列长度相近且相对保守,均具有NINJA 家族所特有的3个结构域:N-端高度保守的LxLxL型EAR基序,139~177位相对保守的NINJA_B结构域及C-端Jas结构域( 图2 )。
为了进一步研究PdMODD与其他 AFP蛋白的亲缘关系,利用MEGA 5.05软件邻接法构建系统进化树,结果( 图3 )显示,PdMODD1与毛果杨PtAFP2亲缘关系最近,并与拟南芥AtAFP1,木薯MeAFP2聚成一个分支;PdMODD2与麻疯树JcAFP2亲缘关系最近,与拟南芥AtAFP2位于一个分支;PdMODD3与毛果杨PtAFP3-1亲缘关系最近,与栓皮槠QsAFP3、拟南芥AtAFP4聚于同一分支。

▲图 2 PdMODD与其他植物 AFP氨基酸序列多重比对
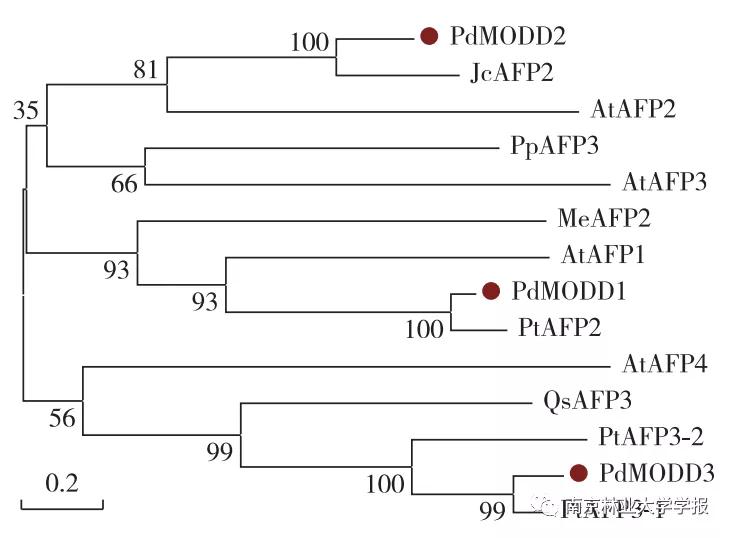
▲图 3 PdMODD 与其他植物 AFP 蛋白系统进化树
3.3 PdMODD 基因亚细胞定位与组织特异性分析
为了研究 PdMODD 的亚细胞定位,将 PdMODD 基因与GFP绿色荧光蛋白基因融合后构建植物融合表达载体pB1121-PdMODD-GFP,通过基因枪瞬时转化,分别将pBI121-PdMODD-GFP质粒转人洋葱表皮细胞,暗培养24 h后在激光共聚焦显微镜下观察 PdMODD 基因的定位情况。结果显示( 图4 ),对照pBI121-GFP( 35S::GFP)在细胞膜和细胞核内都有绿色荧光分布, PdMODD 1- PdMODD 3只在细胞核内有绿色荧光分布,说明 PdMODD 1- PdMODD 3均定位在细胞核中,与预测结果相一致。
通过qRT-PCR分析正常生长条件下 PdMODD 基因在‘渤丰3号’杨不同组织的表达模式,结果显示 PdMODD 1、 PdMODD 2和 PdMODD 3的表达模式相似,均在叶中表达量最高,茎中次之,根中表达量最低,其中 PdMODD 1和 PdMODD 3在茎和叶中的表达量均显著高于根( 图5 ),表明 PdMODD 基因在不同组织间的表达存在差异,具有组织特异性。
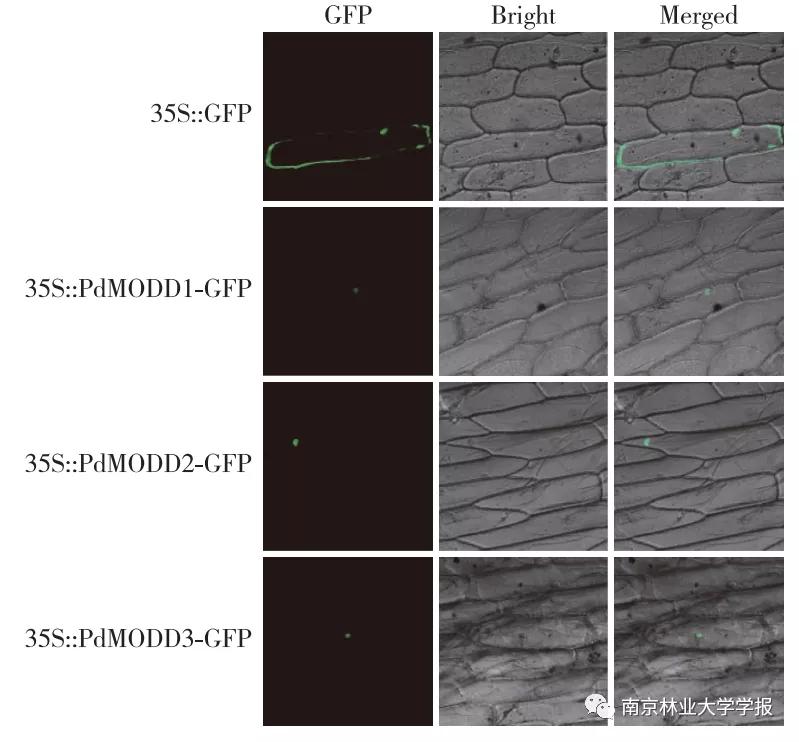
▲图 4 PdMODD 在洋葱表皮中的亚细胞定位
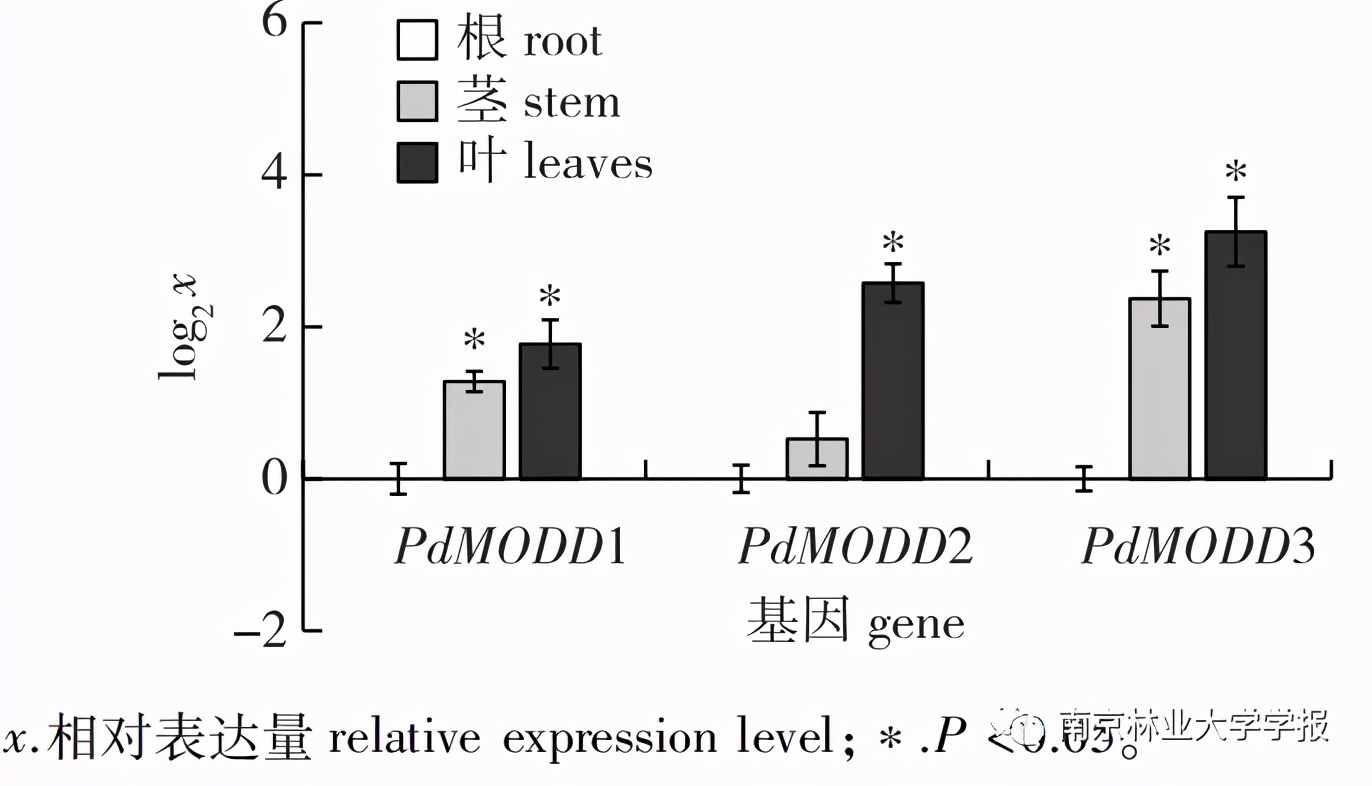
▲图 5 ‘渤丰 3号’杨 PdMODD基因的组织特异性表达
3.4 PdMODD 基因对盐和干旱胁迫的响应
在NaCl和PEG处理条件下,3 h时‘渤丰3号’杨顶湍叶片开始出现萎蔫下垂,随着胁迫时间的推移,叶片萎蔫程度逐渐加重,48 h时萎蔫现象最明显。为了探究 PdMODD 基因与盐和干旱胁迫的应答关系,采用qRT-PCR分析了NaCl和PEG处理下3个基因在根、茎、叶器官的表达模式。结果显示,NaCl胁迫下,根中PdMODD1、PMODD2和PaMODD3的表达趋势基本一致,在3~24 h的表达量均极显著高于0 h(CK),其中 PdMODD 3表达量最高,在3、6、12、24 h的表达量分别高达对照的398.51、454.57、375.30、314.19倍。在茎中, PdMODD 1-3基因均被显著诱导表达,随着胁迫时间增长表达趋势近似于“N”形,表达量均在6 h升高至峰值,分别为对照的38.54、41.46和139.50倍。在叶中,3个基因的表达水平均显著高于对照,且表达趋势随胁迫时间增长呈倒“V”形,均在6 h出现峰值,表达量分别为对照的21.42,16.92、140.31倍( 图6A、6C、6E )。以上结果表明, PdMODD 1、 PdMODD 2和 PdMODD 3基因均能对盐胁迫做出应答。
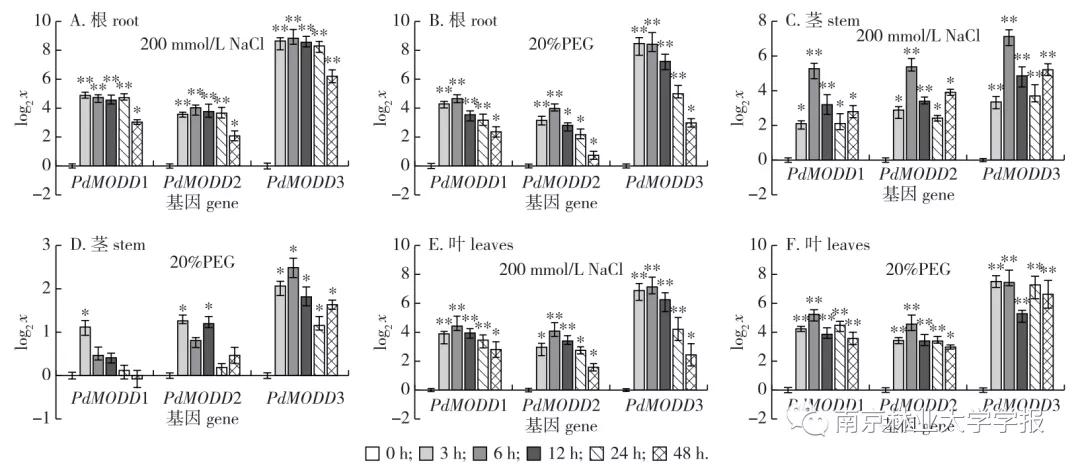
▲图 6 PdMODD基因在 NaCl和PEG胁迫下的相对表达量
PEG胁迫下,根中 PdMODD 1、 PdMODD 2和 PdMODD 3在所有胁迫时间点的表达量均显著高于0 h,且3个基因表达趋势大致相同,近似于倒“V"形,其中 PdMODD 1和 PdMODD 2在6 h表达量最高,分别是对照的25.16和15.70倍; PdMODD 3在3h达峰值,为对照的353.22倍。在茎中, PdMODD 1和 PdMODD 2在大部分胁迫时间点未发生显著变化,而 PdMODD 3在整个胁迫过程中均显著上调表达﹔在叶中,除 PdMODD 2 在 48h 的表达外,3个基因均极显著高于对照,其中 PdMODD 1和 PdMODD 2表达模式基本一致,在6 h表达量最高,分别为对照的37.646和23.56倍, PdMODD 3在3h最高,是对照的181.11倍( 图6B、6D、6F )。结果表明 PdMODD 1、 PdMODD 2和 PdMODD 3可能参与了‘渤丰3号’杨干旱胁迫应答。
4结论
PdMODD 1—3均为NINJA家族成员,含有保守的转录抑制基序EAR,具有组织特异性,在叶中高表达,且能被NaCl和PEG胁迫显著诱导表达,推测 PdMODD 1、 PMODD 2和 PdMODD 3可能会在‘渤丰3号’杨盐和干旱胁追响应中发挥重要的调节作用。

关注我们,更多精彩
