宫颈癌是常见的妇科恶性肿瘤,仅次于乳腺癌。*疆新**是我国宫颈癌高发区之一,在南疆地区维吾尔族妇女宫颈癌患病率和死亡率很高。SiHa细胞株是宫颈癌细胞株研究中应用最广泛的癌细胞。
高良姜素对宫颈癌SiHa细胞凋亡及基因表达的影响
古扎力努尔买提沙1,木塔力甫·艾买提1,艾尔肯·肉孜比拉力2,努尔满古力·肉孜3,
阿布力孜·阿布杜拉1,2
【作者单位】
1.*疆新**地方病分子生物学重点实验室
2.*疆新**医科大学基础医学院生物化学教研室,乌鲁木齐 830011
3.*疆新**维吾尔自治区人民医院妇科,乌鲁木齐 830000
【摘要】目的 探讨高良姜素(Galangin)对宫颈癌SiHa细胞增殖、凋亡及基因表达的影响。方法 从高良姜(Alpiniaofficinarum Hance)中提取分离高良姜素,(1)采用四甲基偶氮唑盐(MTT)比色法,检测空白对照组、不同浓度高良姜素干预组、顺铂(Cisplatin)组SiHa细胞在24、48、72h内的细胞存活率、用荧光倒置显微镜观察对细胞形态的影响。(2)采用流式细胞检测法检测高良姜素在24h内对Siha细胞凋亡的影响。(3)采用荧光定量PCR的方法,分析半胱氨酸的天冬氨酸蛋白水解酶3(Caspase3)和CDK4等基因在高良姜素干预前、后的表达水平。结果 以高良姜素干预宫颈癌SiHa细胞后,与空白对照组比较,细胞存活率明显下降并呈剂量和时间依赖性;早期凋亡率明显上升,并呈剂量依赖性;细胞凋亡因子Caspase3上调表达(P <0.05),呈剂量依赖性,而高良姜素干预前、后细胞周期蛋白激酶CDK4的表达调控无统计学意义(P >0.05)。结论 高良姜素能抑制宫颈癌Siha细胞增殖,促进细胞凋亡,其作用机理可能通过提高细胞凋亡执行因子的活性诱导细胞凋亡,从而表现出抗肿瘤活性,但对细胞周期的影响需要进一步研究。
【关键词】高良姜素;宫颈癌Siha细胞;细胞凋亡;基因表达
【中图分类号】R737.33
【文献标识码】A
【文章编号】1009-5551(2016)12-1595-06
【doi】10.3969/j.issn.1009-5551.2016.12.028
宫颈癌是常见的妇科恶性肿瘤,仅次于乳腺癌[1-2]。*疆新**是我国宫颈癌高发区之一,在南疆地区维吾尔族妇女宫颈癌患病率和死亡率很高。SiHa细胞株是宫颈癌细胞株研究中应用最广泛的癌细胞。本课题组前期研究中,从高良姜提取高良姜总黄酮,并干预宫颈癌SiHa细胞,发现高良姜总黄酮(TFs)能诱导SiHa细胞凋亡,影响细胞凋亡相关基因和肿瘤干细胞标记物表达水平[3-4]。然而,高良姜总黄酮包括高良姜素、槲皮素、槲皮素-3-甲醚、*奈山**酚、*奈山**酚-4-甲醚、二氢高良姜醇和儿茶精等多种黄酮类化合物[5],其抑制作用是上述黄酮类化合物共同发挥作用的结果,各有效成分的抗肿瘤活性和机理待进一步研究。
高良姜素(Galangin,3,5,7-三羟基黄酮)作为一种黄酮类化合物,可从姜科山姜属植物高良姜的根茎、植物的地上部分中提取分离得到。现代药理学研究表明,高良姜素在抗诱变、抗畸变、抗肿瘤、抗氧化和清除自由基、调解代谢酶活性等方面发挥独特的化学防护作用[6-8]。近年来,高良姜素的抗肿瘤活性越来越受到关注,有研究发现高良姜素对CRT 阳性细胞[9]、乳腺癌细胞株(Hs578T、U937[10-11]、人食管鳞癌细胞株(KYSE-510)[6]、肝细胞株[12-13]、TRAIL诱导的前列腺癌细胞[14]、HL-60[15]、人头颈部鳞状细胞癌(HNSCC)[16]、胃癌细胞(SNU-484)[17]、结肠癌细胞(HCT-15、HT-29)[18]、肾癌细胞(Caki-1、786-0)[19]等肿瘤细胞均有诱导凋亡作用,而尚未有文献报道高良姜素对宫颈癌细胞凋亡的影响及机制相关的研究。
本研究首先从高良姜的根部提取高良姜总黄酮,再通过聚丙烯酰胺色谱柱分离方法,从总黄酮中分离高良姜素,进而对宫颈癌SiHa细胞进行药物干预,采用MTT和流式细胞检测技术,检测细胞存活率和细胞凋亡,评价高良姜素的抗宫颈癌SiHa细胞活性。其次,采用RT-PCR 法检测不同浓度高良姜素处理后的细胞中关于细胞凋亡基因半胱氨酸的天冬氨酸蛋白水解酶3(Caspase3)和CDK4表达水平的变化,探讨高良姜素影响宫颈癌SiHa细胞增殖、凋亡和基因表达调控的分子机制,为宫颈癌的临床治疗提供新的药物靶点。
1
材料与方法
1.1 主要试剂与仪器DMEM 培养基、胎牛血清(FBS),双抗(青霉素和链霉素)和胰蛋白酶、四甲基偶氮唑盐(MTT),二甲基*砜亚**(DMSO)和AnnexinV-FITC 等试剂均购自Gibco 公司;Trizol试剂(DP405北京天根生物公司),顺铂(大连美仑),氯仿(上海生工),异丙醇、乙醇(上海生工),聚酰胺(*疆新**恒博鑫业),cDNA 合成试剂盒(Fermentas),SYBRGreen1试剂盒(TAKARA);宫颈癌SiHa细胞由*疆新**地方病分子生物学实验室提供。旋转蒸发仪(Heidolph,德国),CO2 孵育箱(HeraCell-150,ThermoFishers公司),生物安全柜(ThermoKS-18,Thermo Fishers 公司),酶标仪(BeckmannCoulters 公司),流式细胞仪(LSR Ⅱ, BDBiosciences公司),光学显微镜(BA210,Motic公司),定量PCR 仪(Bio-Rad,美国),核酸-蛋白快速测定仪。
1.2 高良姜素的制备将前期研究中按照常规化学提取方法提取的粗提物(高良姜总黄酮)用60mL95%乙醇溶解,慢慢注入到已活化的聚酰胺色谱柱,再用500mL95% 乙醇饱和,过夜;次日分别用3倍柱体积的水及20%、40%、50%、60% 的乙醇溶液洗脱;合并50% 和60% 的乙醇洗脱液,减压浓缩成淡黄色混悬液,放置4℃ 冰箱,静止分层;弃除上层,过滤下层,滤纸上的残渣用无水乙醇收集到表面皿上,静止自然挥发,得黄色粉末状固体,即高良姜素。用DMSO 溶解高良姜素,配置浓度为80mg/mL的母液。
1.3 SiHa 细胞的培养在常规的细胞培养条件下,使用含10% 胎牛血清、100μg/mL 链霉素的DMEM 全培养基培养细胞。当细胞融合度达80%时,用胰蛋白酶(0.25%)消化方法收集细胞,用于MTT、流式细胞检测等后续实验的传代和接种。
1.4 细胞形态学观察和MTT 检测按6×103个细胞/孔密度,将宫颈癌SiHa细胞接种在96孔细胞培养板上,培养。24h后,将96孔板细胞设为空白对照组、不同浓度高良姜素干预组、顺铂组3组,每个组6个复孔。配置空白对照组培养液(200μLDMEM 培养液加入1μLDMSO,即DMSO 浓度为1596 0.5%);配置高良姜素150、125、100、75、50、25μg/mL组干预溶液:将80mg/mL的母液以DMSO 逐步稀释至30、25、20、15、10、5mg/mL 的高良姜素溶液,再按照200μLDMEM 培养液加入1μL药物的比例,将不同浓度高良姜素溶液加入到DMEM培养液中,配置干预浓度为150、125、100、75、50、25μg/mL的细胞培养液;配置顺铂100、50、25μg/mL组溶液:将25mg/mL顺铂母液以DMSO 逐步稀释至20、10、5mg/mL浓度,再用DMEM 培养液稀释至100、50、25μg/mL 浓度,并按24、48、72h 时间梯度对宫颈癌SiHa细胞进行药物干预。药物干预结束后,在24、48、72h时,使用光学显微镜观察细胞形态变化。继而加入5 mg/mL MTT 溶液10μL,孵育4h后,弃去培养基,以150μLDMSO 溶解,在酶标仪上570nm 处读取吸光度并按以下公式计算细胞存活率:细胞存活率= (OD实验组-OD空白对照组)/OD空白对照组×100%。
1.5 细胞凋亡检测在6孔培养板上按1×105个细胞/孔密度培养细胞并进行药物干预;24h后收集细胞(约3×104个细胞),依次加入400μL细胞凋亡检测试剂盒提供的结合缓冲液(1×bindingbuffer)、5μLAnnexinV-FITC和10μLPI,在室温下避光反应15min,在流式细胞仪上检测细胞凋亡率。
1.6 RT-PCR 检测细胞凋亡相关基因mRNA 表达水平以25μg/mL 和100μg/mL 高良姜素干预细胞24h 后,收集细胞,用Trizol试剂提取总RNA,利用核酸-蛋白快速测定仪测定所提取RNA浓度和纯度,用1% 琼脂糖凝胶电泳对RNA 进行质量检测。随后抽取1μg总RNA 利用逆转录试剂盒并按说明书合成cDNA;再以cDNA 为模板,并以β-actin为内参照,相应基因上下游引物(表1)及SYBR对cDNA 进行扩增。RT-PCR 反应条件为:DNA 变性95℃3min,合成40个循环(95℃10s,55℃~62.6℃30s)。
1.7 统计学处理以SPSS17.0版专用统计分析软件对数据进行一般线性模型单变量方差分析,实验数据以均数±标准偏差(-x ±s)表示,以P <0.05为差异有统计学意义。通过iQ5软件得到候选基因在样品中的相对表达水平。以β-actin为内参基因,计算基因表达水平,表达水平(相对表达量)=目的基因的起始量/内参基因的起始量。
2
结果
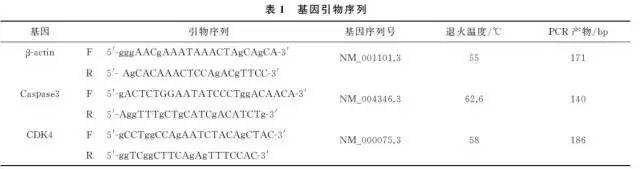
2.1 高良姜素对SiHa细胞形态的影响在24h,与空白对照组相比,随着高良姜素和顺铂浓度的增加,细胞数量明显减少,细胞胞体缩小,皱缩或碎裂,培养液中有较多的细胞碎片,悬浮细胞增多(图1、2);在观察不同时间段细胞状态时,发现药物干预后细胞增殖比正常细胞增殖缓慢(图3)。
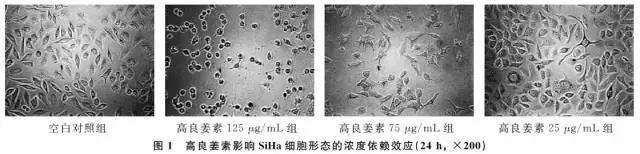
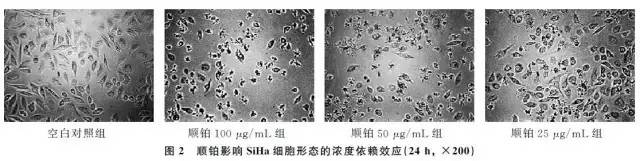
2.2 高良姜素干预对SiHa宫颈癌细胞存活率的影响与空白对照组比较,随着药物剂量增加和干预时间延长,SiHa细胞增殖活力(存活率)均梯度性下降,差异有统计学意义(P <0.05),呈剂量和时间依赖性;与顺铂组(Cisplatin,DDP)相比较,其效应基本相似(表2)。
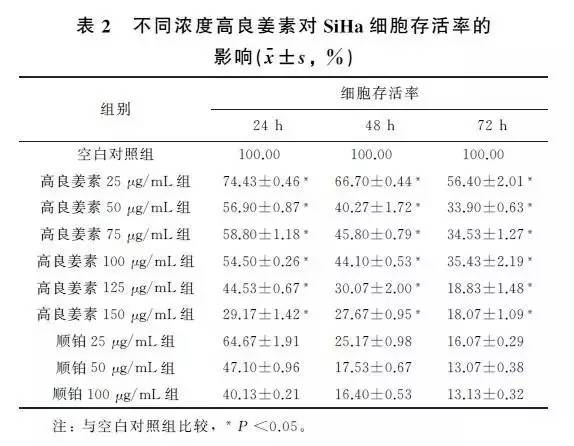
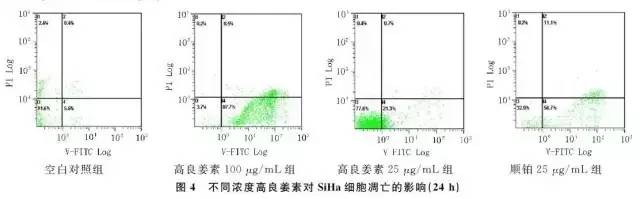
2.3 高良姜素对SiHa宫颈癌细胞凋亡的影响在空白对照组,SiHa细胞自然发生的早期和晚期凋亡指数分别为5.6% 和0.4%,以25、100μg/mL 高良姜素干预24h 后,早期凋亡率分别为21.3% 和87.7%,晚期凋亡率分别为0.7% 和8.5%;与空白对照组比较,早期凋亡率明显上升,呈剂量依赖性。以25μg/mL 顺铂干预24h后发生的早期凋亡和晚期凋亡率分别为56.7% 和11.1%(图4)。
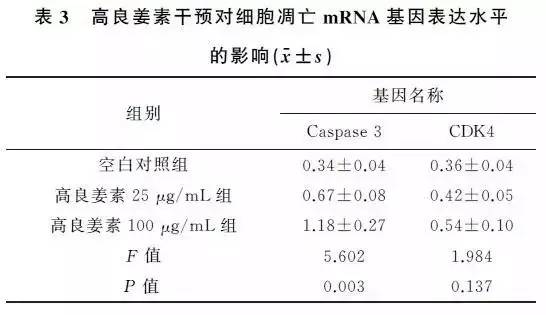
2.4 高良姜素干预对细胞凋亡相关基因表达水平的影响与空白对照组比较,25μg/mL和100μg/mL的高良姜素干预24h后,SiHa细胞内Caspase3基因的mRNA表达水平上调,差异有统计学意义(P <0.05)。高良姜素100μg/mL组Caspase3的表达水平高于高良姜素25μg/mL组,差异有统计学意义(P<0.05)。高良姜素干预后CDK4的表达水平上调,与空白对照组比较,差异无统计学意义(P >0.05),高良姜素25μg/mL组及100μg/mL组CDK4mRNA表达水平差异无统计学意义(P >0.05)(表3)。
3
讨论
宫颈癌的发病机制主要与病毒感染[21]、癌基因与抑癌基因、细胞周期调节、端粒与端粒酶等有关。其中,人乳头状瘤病毒(HPV)的单一或多重持续感染现已被公认为是宫颈癌发生、发展的关键因素[22]。95% 以上的宫颈癌是由高危型HPV 引起的,其中HPV16与宫颈鳞状细胞癌的发生密切相关,HPV18则与宫颈腺癌相关[23]。近年来研究发现黄酮类化合物在治疗肿瘤、心血管疾病、糖尿病、炎症、细菌、病毒感染(抗HIV 病毒等)等多方面具有药理作用,而上述作用归因于其特有的化学结构,即黄酮结构中一定数目的羟基(2~4个)、C环2、3位的双键、2位的β环定位、3位和6位的羟基、β环上邻位的羟基是发挥抗肿瘤活性的结构效应元件。诸多研究表明,黄酮类化合物主要是通过阻滞癌细胞增殖、诱导癌细胞凋亡、抑制蛋白激酶活性、抑制信号转导途径等机制发挥抗癌效应[24]。本研究中使用的高良姜素是从中药高良姜的根部提取纯化的单体黄酮类化合物,其分子结构中C环2、3位的有2个双键,2位的β环定位、3位有羟基(即β环上邻位的羟基),符合上述黄酮类化合物发挥抗肿瘤作用的结构特点。本课题组在前期研究中,通过细胞水平研究,发现高良姜总黄酮能够抑制肿瘤细胞增殖,诱导细胞凋亡[3]。本研究在此基础上,进一步对高良姜总黄酮中高良姜素(单体)的抗癌活性进行研究。本课组前期研究报道了以25~150μg/mL浓度的高良姜总黄酮干预SiHa细胞后,其细胞存活率分别为86.26%、80.05%、60.29%、59.51%[3]。
本研究中,在25~150μg/mL浓度范围的高良姜素干预后的细胞存活率均明显低于高良姜总黄酮干预后的细胞细胞存活率。MTT 和细胞凋亡结果显示,高良姜素抑制宫颈癌Siha细胞增殖,降低细胞存活率,促进细胞凋亡,呈剂量和时间依赖性。因此,根据实验结果和上述结构分析可推测,高良姜素具有抗宫颈癌活性。
Caspase是半胱氨酸依赖性细胞死亡蛋白酶,是一大类凋亡调节基因。Caspase3是凋亡执行因子,其能够直接降解胞内的结构蛋白和功能蛋白而引起凋亡[25]。CDK4 (cyclinDdependentkinase4)是一种细胞周期蛋白激酶,在p16 (INK4A)-CDK4/6-cyclinD1-pR-Rb-E2F信号转导通路中可与cyclinD1形成复合体,抑制pRB 活性,促进肿瘤细胞增殖、分化[26]。本研究采用RT-PCR 方法,进一步研究高良姜素促进癌细胞凋亡的分子机制,研究结果显示,高良姜素通过提高凋亡执行因子Caspase3的转录水平、促进细胞内结构蛋白的降解,实现凋亡作用。因此,高良姜素可能是高良姜总黄酮中发挥主要作用的有效成分,这对基于高良姜素的抗肿瘤新药研发有重要意义。
本研究中发现药物干预后CDK4基因的转录水平没有明显变化,而在前期研究中,吾尼且木·吐拉克等[4]发现高良姜总黄酮干预宫颈癌Siha细胞使CDK4基因下调表达,说明高良姜总黄酮中可能有另外一种单体黄酮类药物影响其表达水平,这部分内容需要进一步研究。另外,在以往研究中报道,干细胞标记物是胚胎干细胞阶段的转录因子,在多种肿瘤干细胞中过度表达,可能与肿瘤组织的形成、侵袭转移关系密切[27-29]。在今后的研究中进一步探讨良姜素与宫颈癌干细胞信号转导网络的相互作用机制,是新一代抗宫颈癌药物研发的重要切入点。
【参考文献:略】