SARA M. LEWIS , CHOONG HAY WONG, AVALON C.S. OWENS , CANDACE FALLON, SARINA JEPSEN,ANCHANA THANCHAROEN, CHIAHSIUNG WU, RAPHAEL DE COCK, MARTIN NOVÁK, TANIA LÓPEZ-PALAFOX, VERONICA KHOO, AND J. MICHAEL REED
作者:Sara Lewis 翻译:萍曦 校对:吴加雄 全文读完约需12分钟
关键词: 鞘翅目、灭绝危机、昆虫保护、IUCN、萤科
自它们进化了大约2.97亿年以来(Zhang et al. 2018),甲虫已非常成功;它们占了已知昆虫种类的38%(Stork 2018)。而萤火虫(鞘翅目:萤科)跻身于极赋魅力的甲虫之列,独特的生物发光求爱行为使他们成为昆虫保护中,一支蛰伏已久的旗舰物种。
萤火虫在全球有超过2000种物种,令人惊讶的是,其展现了多彩的生命历史特征。包括在白天活动时段的不发光的成虫,能发光却不能飞翔的雌性萤火虫,能发光的萤火虫通过交换特定的闪光信号来交流。萤火虫还栖息在生态多样化的栖息地中,包括湿地(例如红树林,稻田,沼泽,沙漠渗水),草原,森林,农田和城市公园。
萤火虫幼虫有可能是水生,半水生或者陆生的,在那数月,甚至数年的时间都以蜗牛,蚯蚓和其他软体动物为食。相反,萤火虫成虫通常寿命短且不觅食。某些萤火虫分类群,对于食饵或是栖息地高度专一,而有些萤火虫分类群則适应性相当广(Reed等人,2020年)。
萤火虫在许多国家而言,具有重要的经济意义,因为它们代表着日益增长的生态旅游中的亮点(Napompeth2009,Lewis2016)。然而,正如许多无脊椎动物一样(Cardoso 等,2011),萤火虫在全球保护行动行列中被严重忽视了。
对于几乎所有的萤火虫种类来说,种群变化趋势的定量监测数据的研究十分缺乏。然而,调查显示近期马来西亚的红树林萤火虫(种类)和英国的萤火虫(Lampyris noctiluca ) (Gardiner 2011, Atkins et al. 2017)数量显著下降。
轶事报告和专家意见还表明:在近几十年中,诸多萤火虫种类出现的频率和数量都减少了(Lewis 2016,Faust 2017,Lloyd 2018)。在2010年,一个国际萤火虫专家组织在马来西亚召集,并撰写了《雪兰莪州萤火虫保护宣言》, 建议采取行动来保护这些独特的昆虫。
2018年,IUCN(自然保护世界国际联盟)萤火虫专家小组成立,旨在评估全球萤火虫保护状况和灭绝风险。作为这项工作的一部分,在本文中我们将根据来自不同地理区域的专家的意见调查,讨论对萤火虫种群多样性及稳定性的潜在威胁。我们也将审查有关此类对萤火虫族群构成威胁的影响因素。最后,因为这些特定物种的行为和生命史特征也使得它们在遭受到各种风险时,承受度不一而同。所以对于每一种威胁,我们将针对其造成的威胁相关度逐一进行讨论。
萤火虫专家的全球调研

图1.萤火虫(鞘翅目:萤科)在生态、行为或灭绝风险方面都各不相同。 (a)Photinus pyralis 的雌性和雄性成虫都具有飞行能力,其种群纵横整个美国东部,但基因流动性仍十分有限。(照片:Terry Priest)(b)例如其雌成虫不会飞行的雌光萤这类种群的分布更为局限。(照片:Zdeněk Chalupa)(c)所有萤火虫幼虫都是掠食性昆虫,且它们的食饵单一;L. noctiluca(一种萤火虫)正出攻击Helin aspersa(一种蜗牛)。(照片:Heinz Albers)
(d)萤火虫大量汇集、同步发光求爱的行为形成一道别样的风景从而带动了生态旅游,然而这也可能导致当地的生境退化。(Pteroptyx malaccae in Thailand, 照片: Radim Schreiber)
萤火虫国际联盟由全球对萤火虫感兴趣并致力于萤火虫生态学、行为学、分类学或者萤火虫保护的个体组成的科学组织。2019年1月,我们通过邮件向350名萤火虫国际网络联盟中成员发送了一个简短的问卷调查。这些调查结果应该被重视起来,因为它们反映的专家意见,仅仅只是对于萤火虫种群稳定性的感知性威胁。
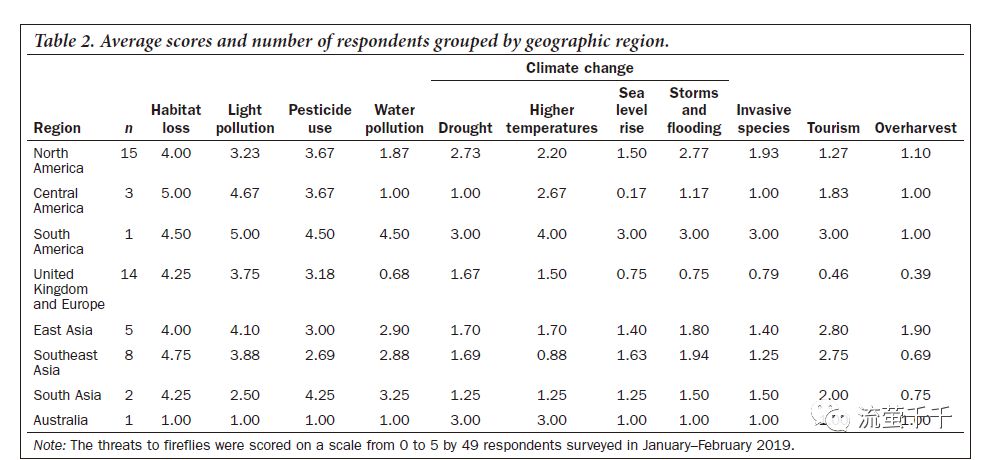
我们将受访者分为8个地理区域:北美(美国、加拿大)、中美洲及墨西哥、亚洲南部(印度、斯里兰卡),东亚(中国台湾、中国香港、中国、日本),和东南亚(泰国、马来西亚)。我们分别有一个受访者来自澳大利亚和南美洲,而非洲则没有。全文仅使用提供所有威胁评分(n = 49)的受访者数据,我们就可以将调查结果显示为八个地区和全球范围内全球威胁评分的平均值。
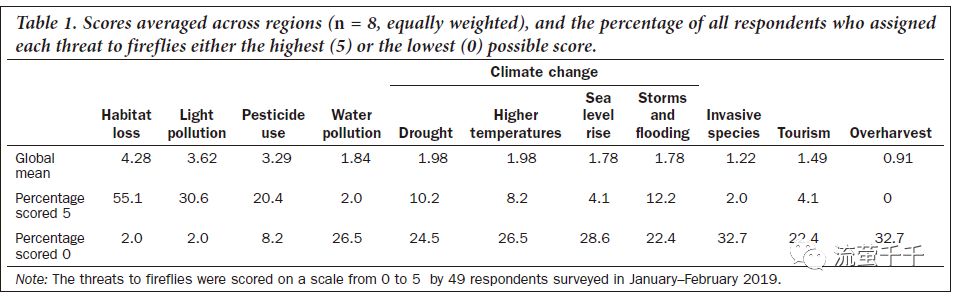
受访者对于特定威胁所得出的观点,总结如下(获取详细的受访者观点,可查阅补充的材料)。我们还通过文献去寻找关于这些潜在的威胁如何影响萤火虫的生存,繁衍或者影响种群平衡的现有证据。不止于此,还有受访者提到的其余的一些威胁(Box 1)

调查结果
01
栖息地的破碎化与消失
生境丧失被认为是全球乃至于所有区域的萤火虫最严峻的威胁。生境研究者认为,生境的丧失和碎片化预计为是显著问题(Reed et al.2020)。对于在美国东部广泛分布且数量较多的Photinus pyralis,在种群间进行全基因组SNP(单核苷酸多态性)分析得出平均Fst(固定指数)值为0.38(Lower et al.2018)。如果这一结果适用于其他萤火虫种类,那么这种程度的遗传隔离意味着灭绝的种群不太可能通过迁徙来挽救。
不会飞翔的雌性种群的扩散距离更为局限,这里其中包括 Lampyris noctiluca (Atkins et al. 2017) and Phosphaneus hemipterus; 在后者中,雌雄都不能飞(De Cock 2009)。幼虫爬动造成形成的传播距离也比较有限:在整个幼虫时期,Luciola par- vula的陆生幼虫预计其的扩散距离也仅仅只有几米。(Kakehashi et al. 2014)。然而,扩散距离对于水生幼虫来说可能更高,因为它们可能会沿着河流和灌溉渠,就像其他无脊椎动物被报道的那样 (e.g., Perry and Perry 1986)。
在泰国,水生萤火虫Sclerotia aquatilis的卵和幼虫常附在浮萍上,所以可能随着这些水生植物一起漂流。如果萤火虫雌虫和陆生幼虫被洪水带走并有一定的数量幸存下来,他们可能从此在这一片适合生存的地方繁衍生息。
在欧洲,萤火虫的栖息地已因城市化,工业化和农业集约化而丧失(De Cock 2009)。农业集约化——意味着生境丧失和碎片化,同时也增加了杀虫剂,除草剂和化肥的使用——已经被公认为是大量昆虫种群数量下降的一个重要的推手(Wagner 2018, 2020).。在整个英国,由于放牧地被抛弃,萤火虫(Lampyris noctiluca)经常出没的草原生境已因农业集约化和林地演替而丧失(Gardiner 2011)。长期调查英格兰南部和中部几个地点的夜蛾L.noctiluca记录到人口大量减少,这可能是由于土地利用的变化(例如,道路建设,沟渠填充,木材堆放)以及干燥(Gardiner 2011, Atkins et al. 2017)。在意大利,一位调查受访者认为农业集约化导致帕达纳平原和北部Appenines上的意大利Luciola italica,Luciola lusitanica和Lampyris萤火虫数量减少。
相反,另一个在西班牙地中海起地区的受访者表示对废弃小果园和灌溉农业用地担忧,Nyctophila reichii, Lampyris iberica, 和Lamprohiza paulinoi经常出现。一旦被抛弃,这些曾是耕种的地方变得更加干燥、不适合蜗牛生存,而蜗牛恰恰是某些萤火虫的主要猎物。
在日本,面对发展和农村人口外迁,一个以山岳而闻名的标志性传统景观正在消失(Kobori and Primack 2003)。这个管理有序的栖息地由溪流,池塘,稻田和被森林包围的耕地组成的乡村组成,曾经为日本乡村包括萤火虫的大量生物多样性提供了支持(Oba等,2011)。在马来西亚,河岸边的红树林逐步被农业,水产养殖和城市化侵蚀,导致生活在这里的 Pteroptyx tener萤火虫繁衍群体递减 (Jusoh and Hashim 2012, Khoo et al. 2014)。在整个东南亚地区,大片的河岸红树林的区域已被清除,以用于油棕种植,虾场或减洪,使得这些区域不适合 Pteroptyx萤火虫幼虫及其食物蜗牛的繁衍(Wong 2009, Nada et al. 2009, Thancharoen 2012, Wong and Yeap 2012, Jusoh and Hashim 2012, Jusoh et al. 2010, Khoo et al. 2012, 2014)。夜里,Pteroptyx成虫常聚集在沿岸地红树林上,用光来交流彼此的爱意,然而这些树现已被大量地移除了(例如,图3a)。
巴西的大西洋雨林具有很高的萤火虫多样性(Viviani 2001, Viviani and Santos 2012, Silveira and Mermudes 2013, 2014),却也是世界上受威胁和破碎程度最高的雨林之一(Hoorn et al. 2010)。在墨西哥的特拉斯卡拉(Tlaxcala),Macroplampis palaciosi(另一种雌性萤火虫成虫也不能飞行的种类)种群残存于因大量砍伐而破碎的森林碎片中(Vance和Kuri 2017)。在北美东部,萤火虫栖息地的流失主要是因为城市化以及房地产开发造成的(例如,图3b)。
在全球范围内,沿海岸线的人口增长已导致广泛的栖息地丧失和破碎化 (Polidoro et al. 2010),同时也威胁着红树林萤火虫和其它栖息于沿海湿地的物种。
在特拉华州有一个 十分特殊的海洋栖息地Photuris bethaniensis , 它被发现于海边沙丘之间的淡水洼地中(Heckscher,2010年)。其湿地生境面临着大规模住宅开发,其威胁迫在眉睫(Kitt Heckscher, Delaware State University, Dover, Delaware, personal communication, 2018)。
在佛罗里达州,Micronaspis floridana 栖息在完整的沿海红树林和盐水湿地中;现在该物种的分布受到很大限制(Faust 2017)。在美国西部和德克萨斯州,一些萤火虫在毗邻永久水源的栖息地(河流,溪流,湖泊,池塘,泉水和灌溉农田)里夹缝生存。抽取地下水供应城市和农业用水,导致地表水流量减少、地下水位降低 (Larry Buschman, Kansas State University, Lawrence, Kansas, personal com- munication, 2019),同时气候变化导致了干旱的增加,可能会进一步压缩萤火虫在这片区域适合栖息的家。
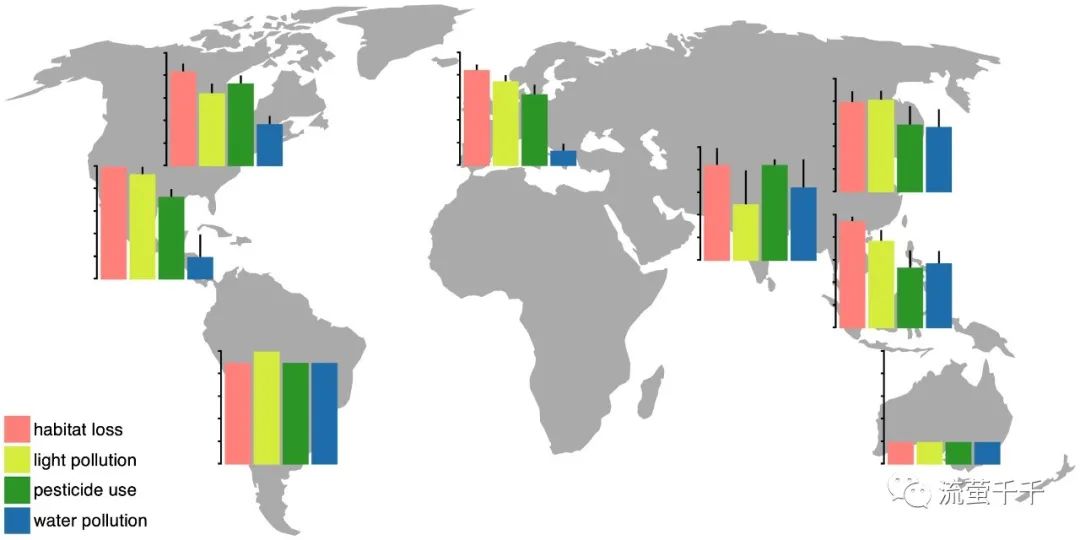
图2.萤火虫四大威胁指数地理分布(条形图表示平均值+一个标准误差(0-5),整理自2019年1月至2月接受调查的49位各地学者的报告。其他威胁的指数的样本量及所在地分布,参见表二。
02
夜间的人造光污染(ALAN)
在全球范围内,夜间人造光(ALAN)被认为是萤火虫的 第二大严峻威胁(表 1)。ALAN 包括影响局部区域的直向照明(例如,石化厂的煤气火炬,路灯,运动场,商 业标牌,安全灯,广告牌)、天光和漫射照明(其甚至可以超过满月时月光的光亮水平,可以将光亮散射到远离城市中心之外的地方)。
据保守估计,目前全球已有23%的陆地面积被一定程度的ALAN覆盖 (Falchi et al. 2016)。在东亚和南美,光污染被认为是萤火虫的最大威胁,而在大多数其他地区则是第二或第三大威胁(表2)。
ALAN被推测为对于夜行性萤火虫类群的影响尤为突出,毕竟这类成虫依靠生物发光的求爱信号来寻找伴侣(Lloyd 2008,Lewis 2009)。观察和实验研究提供的证据,表明 夜间人造光会对萤火虫种群产生不利影响(有关综述,请参见 Owens 和 Lewis 2018;还有 Mbugua 等人 2020)。
多项研究表明,高水平的ALAN与萤火虫丰度是负相关关系。例如,调查显示,意大利都灵市灯火通明的区域,Luciola italica种群集体缺席 (Picchi et al. 2013)。同样,在巴西圣保罗,多种萤火虫也被仅在夜间人造光含量较低的地区生存( Hagen 和 Viviani 2009 , Viviani 等人 2010)。因为夜间人造光 与城市化息息相关。
然而,这些调查研究难以分离出萤火虫种群减少的主因。在这种情况下,各地的信息是不可缺的补充来源;一位调查对象指出,中小村庄周围光污染的增加是造成西班牙萤火虫减少的原因。Owens和Lewis(2018)审查了实验研究,证明了人造光会干扰萤火虫求爱信号的产生与接收。雌性Lampyris noctiluca会产生持久的光亮,以吸引飞行中的雄性,多个野外的调查发现,各种类型的ALAN都会降低雄性的吸引力(Ineichen和Rüttimann 2012, Bird和 Parker 2014, Bek 2015)。
许多发光的萤火虫加入到“光语”中,在这神秘的“光语”中,雌性会对雄性求爱,予以闪光回应(Lloyd 2008)。通过对三种萤火虫类群(Pteroptyx maipo(Yiu 2012),Photuris(Firebaugh和Haynes 2016)和雄性Photinus(Hagen et al.2015,Costin和Boulton 2016)的实验,发现是人造光的干扰减少了萤火虫的求偶发光行为。此外,暴露在人造光的雌性Photinus pyralis对雄性求爱信号的响应频率降低(Firebaugh and Haynes 2016),交配成功率无明显减少。因此,多个证据表明夜间人造光干扰了萤火虫的繁殖行为,并可能增加灭绝的风险。

图3.全球萤火虫面临的最大威胁是栖息地的丧失和夜间的人工照明。(a)在马来西亚,清雪兰莪河沿岸的红树林被清理,以便于农业和水产养殖,致使萤火虫成虫的求偶地盘、幼虫的栖息环境被破坏(照片:Laurence Kirton)。(b)在特拉华州,住宅开发威胁到仅生存于淡水里的 sw的Photuris bethaniensis 种群减少。(c)基于2006年全球卫星数据的ALAN地图,颜色表示人工对自然天空亮度的比率:黄色表示亮度提高1至3倍,橙色代表亮度提高3至9倍,红色代表亮度提高9至27倍。David Lorenz的地图,https://djlorenz.github.io/astronomy/lp2006。
03
农药的使用
在全球范围内,农药被认为是萤火虫的第三大严重威胁(表1),地理区域之间有些差异(表 2)。常见的农业杀虫剂包括各种有机氯, 有机磷酸盐,以及最近出现的新*碱烟**类 (Simon-Delso et al. 2015)。尽管只有很少的研究调查了它们对萤火虫的直接影响(见下文),但已知这种广谱杀虫剂会对许多非目标昆虫和其他类群产生不利影响(reviewed by Sanchez-Bayo 2011, Pisa et al. 2015)。
萤火虫可能被空中喷洒的农药中 头奖,也可能接触到含杀虫剂的土壤或水、又或者吃了被污染的猎物。对于萤火虫而言,水和土壤中高浓度的杀虫剂可能对其伤害性十分大。因为幼虫阶段要么在水下(例如水生萤火虫,如 Aquatica 和 Sclerotia),要么在河岸红树林的根部,或在成虫求爱时所停歇树木背后的植被中 (Pteroptyx fireflies),或者在泥土里(例如,陆生萤火虫,如Lampyris,Photinus和Photuris)杀虫剂也可能会侵扰萤火虫其它的生活阶段,因为其卵产在土壤,苔藓或腐烂的木头中,而蛹在地下或树干上发育。当成虫在撒过农药的土壤或树叶上休息时,也就暴露在了杀虫剂残留物中。
迄今为止,仅在两项公开的实验室研究中测试了直接接触农药对萤火虫的影响。根据Tabaru及其同事1970 年的报告陈述,尽管含有5%的硫磷的浓缩乳化型杀虫剂对源氏萤的幼虫及其猎物蜗牛都具有毒性,但当以 5%-10%的可湿性粉剂的形态进行测试时,有机磷杀虫剂(杀螟松,二硫磷和二苯硫磷)对二者仍呈低毒性。在另一项研究中,是利用韩国水生的平家萤进行的,在厂商推荐的浓度下测试了10种杀虫剂,发现暴露于9种化合物中的幼虫和成虫的死亡率均增至80%–100%,而暴露于8种化合物中的情况下,卵的孵化率降至0%–33Lee et al. 2008)。虫酰肼)致使幼虫死亡率为33%,而成虫死亡高达73%,但并未降低卵的孵化率。而氟虫腈可致使83%的幼虫死亡,成虫的死亡率达100%,卵孵化率降至67%。
在东南亚,油棕种植园和虾场的农业径流危害着幼虫时期的水生或半水生萤火虫。2002年至2003年在马来西亚雪兰莪河进行的一项研究发现,几种农药的含量有时超过了淡水生物的可接受的极限(Leong et al. 2007).。在日本,河流的工业污染和农药污染导致了二十世纪下半叶发现的源氏萤和Aquatica laterali的数量下降(Yuma 1993,Ohba 2004)。
尽管欧盟于2018年4月禁止在室外使用新*碱烟**类药物,但在包括美国在内的其他地区,这些化合物仍在农业和住宅环境中广泛使用(Bonmatin et al.2015)。
在美国,几乎所有玉米和大豆种子都被常规性地涂上新*碱烟**类杀虫剂(Douglas and Tooker 2015),也致使这种农药持久地残留在大片的土壤中。在单个生长季节进行的田间试验中,经可比丁素处理过的玉米种子的田地与对照田相比,萤火虫成虫的丰度降低70.4%(Disque et al.2018),这很可能是由于在经农药处理过的土壤中,幼虫死亡率更高(Galen Dively, University of Maryland, College Park, Maryland, personal communication, 2019)。吡虫啉是常出售给屋主杀死白蛆(金龟子的幼虫)的新*碱烟**类化学物;在一项为期3年的研究中,这种农药在草坪上的应用大大减少了非目标昆虫的数量,而非目标甲虫的数量减少了2.4倍(Peck 2009)。
杀虫剂(如拟除虫菊酯)被广泛用于控制蚊子成虫,但也可能影响非目标昆虫(Davis等,2007)。萤火虫尤其容易受到威胁,因为通常在黄昏时,即蚊子和萤火虫都活跃时才进行喷洒。在对有益的瓢虫Harmonia congengens进行的生物测定现场中发现:只需超少量的苄氯菊酯,就可以使接触到药物的甲虫呈现高死亡率(Peterson et al. 2016)。农药也可以通过削弱幼虫的行动能力或者增加其食物(包括蜗牛和蚯蚓等)的毒性,间接地影响萤火虫 吡虫啉和其他新*碱烟**类化合物已被证明对蚯蚓具有高毒性(有关综述,请参见Sanchez-Bayo 2011,Pisa等人,2015),而它们恰恰是北美萤火虫幼虫的主要猎物(Lewis 2016)。这些猎物其体内累积新*碱烟**类物质(Douglas and Tooker 2015,Chevillot et al.2017),这对于萤火虫幼虫来说,多了一条接触农药的途径。
最后,为了对付各式各样的生物防控对象(包括包括真菌Metrahizium和Beauvaria bassiana和Steinernema sp.线虫),萤火虫作为非靶向生物也跟着糟了秧。(2017年8月;请参阅补充材料)。
04
其它威胁
通过22%–33%受访者判断,在我们的调查中,其余威胁的平均得分很低,可将威胁指数得分视为为0(表示无威胁)。但是,某些特定地区的受访者特别强调了水污染对萤火虫构成潜在威胁。遍及亚洲和南美地区的受访者认为:含化肥,农药的农业废水、工业废水和其他污水的径流是萤火虫的第三或第四大威胁(表2)。与陆生萤火虫幼虫相反,许多亚洲水生萤火虫的幼虫栖息在淡水池塘、河流和溪流中,觅食成长,也经历着虫生的几个幼龄阶段(Lloyd 2008)。这个阶段通常持续几个月,在此期间,幼虫及其猎物蜗牛都将暴露在污水中。
05
旅游
萤火虫旅游已在日本、马来西亚和中国台湾流行开来已久,近来在其他国家(包括泰国(Thancharoen 2012),美国(Faust 2009, 2017)和墨西哥(Vance and Kuri 2017)类似的娱乐活动激增。
经统计,萤火虫旅游吸引了每年超过20万游客(Lewis 2016),给当地带来了可观的经济收入。那么如果此种旅游业疏于管理,则会干扰萤火虫栖息地或影响了成虫繁殖,从而对当地的萤火虫种群生存繁衍构成威胁。
尽管许多萤火虫种类,其生物发光景象备具吸引力,但大多数是处于威胁中的是那些自主同步发光的萤火虫。以美国的Photinus carolinus为例,这类萤火虫的雄性,成百上千地聚集在一起有节奏性地同步发光、闪烁,无不让人惊叹万分。在东南亚,同步闪烁的萤火虫在潮汐河沿岸的红树林中大量地聚集在一起,成为当地重要地旅游亮点(Wong andYeap 2012)。
商业旅行设施、手电筒甚至照相机闪光灯带来了ALAN(Thancharoen和 Masoh 2019)可能会干扰Pteropytx(某种萤火虫)的求爱行为。在泰国,沿红树林线的河流,因Pteropytx旅游业的发展,高速汽艇随之而来致使河岸被侵蚀、树木倒塌,摧毁了幼虫的家园。当其它不会飞的雌性从地面或低矮的植被中发出信号时,却不小心遭游客踩踏。其中包括北卡罗莱纳州的Phausis reticulata和墨西哥纳纳卡米尔帕的Macrolampis palaciosi。
06
过度捕捉
尽管目前尚未被视为威胁,但从历史上看,野生种群大量捕捉萤火虫可能会导致某些种群减少(Bauer等人,2013)。在19世纪末20世纪初,日本源氏萤经历了商业性地捕捞(Lewis,2017年)。
在美国,从1960年至1995年左右,西格玛化学公司每年捕捞约300万北美萤火虫,以提取萤光素酶和萤光素(发光化合物;Lewis 2016)。
在中国,从2009年至2017年期间,数百万计地萤火虫被捕捉,销售给主题公园展览或被包装成浪漫礼物卖给城里人。(Lewis and Owens 2017)。由于萤火虫保护组织持续展开的抗议和*访信**战役,中国野生萤火虫的商业性捕捉才告一段落(LeiPing, Firefly Ecological Alliance, Guangzhou, China, personal communication, 2019)。
07
入侵物种
大多数受访者没有将外来入侵物种定为主要威胁,但红火蚁在美国南部扩散的情势下,萤火虫的幼虫仍在地面上活跃,例如Pyractomena borealis,这很可能对其构成一个新的威胁。
08
气候变化
尽管人为的气候破坏对萤火虫种群的影响仍然未知,但是某些萤火虫对活动范围有限并对栖息环境有着特殊的要求,这也表明它们可能受到干旱和海平面上升的威胁(Reed et al. 2020)。萤火虫在其整个生命阶段都需要潮湿的条件(Lloyd 2008,Atkins等,2017,Evans等,2018),因此干旱持续时间、频率或强度的增加(可能与积雪减少有关),可能使一些地区的萤火虫种群灭绝。(但请参见补充表S1; Harris et al. 2019)。比如说,干旱的热带山地地带的扩张(Wagner 2018,Janzen和Hallwachs 2019)可能威胁着生活在潮湿的新热带森林中的萤火虫类群。同样地,在澳大利亚和美国西部,萤火虫仅存于永久水源中。而干旱,要么直接导致萤火虫死亡,要么萤火虫幼虫因食物短缺而一命呜呼。
在马里兰州和特拉华州,Photuris salina栖息在于沿海微咸和盐沼的较干燥部分,其栖息地容易因海平面上升而被淹没(Heckscher 2010)。如上所述,特拉华沿海物种P. bethaniensis也同样易受害,因为它占据了在大西洋500米以内的沿海沙丘之间淡水洼地,且仅低于当前海平面0.5米(Heckscher and Bartlett 2004)。
在东南亚,Pteropytx的几种萤火虫都聚集在高大的,抢眼的红树林中,这种树的耐盐性较低(Nada等,2009),可能会遭受海水入侵的威胁。
Moving forward
这篇文章以全球视角分析了萤火虫种群的下降甚至灭绝的严峻威胁,我们的调查结果阐明了那些学识渊博的受访者认为这是对于萤火虫物种来说,造成的持久性最重要威胁,也揭示了各个地理区域之间的差异。尽管这些观点显然不能用来确定此类威胁的利害或威胁程度,但我们的结果与其它对昆虫丰度和生样性普遍下降的多方面原因的评估相一致(e.g., Leather 2018, Wagner 2018, 2020, Homburg et al. 2019).。我们相信这些信息有利于将来研究影响萤火虫多样性和丰度的因素。
此外,我们的文献综述提供了有关此类威胁是否及如何影响萤火虫种群的现有证据进行了全面小结,并描述了可能增加某些萤火虫物种对特定威胁的敏感度的风险。这个观点还强调了需要投资在监测研究的紧迫性,这些研究可以提供长期数据来追踪高危萤火虫物种和栖息地的丰度和多样性趋势。除几个显著例外外,有关萤火虫种群趋势的大多证据都是传闻,需要开展来制定一系列标准化的监测方案。此外,还需要进行实验研究来判定在萤火虫生命阶段中,常见杀虫剂对其造成的急性中毒和慢性中毒。
行动起来
基于对觉察威胁的调查,我们对威胁萤火虫的现有证据进行了审查,为保护这些迷人的小精灵,我们制作了如下的行动建议:
保护合适的栖息地
我们需要确定濒临灭绝的物种,并于濒危萤火虫的场地建立避难所。这样做时,必须考虑每一个生命阶段对栖息地的需求,从而确保为萤火虫幼虫及其猎物、化蛹场所、成虫求爱展示和和雌虫产卵提供合适的栖息地。
萤火虫有可能成为建立关键生物多样性地区的旗舰物种。
在马来西亚,河岸红树林和岸边的土地迅速流失对既有经济价值的生态旅游亮点的Pteroptyx的多种萤火虫构成了持续威胁。因此,划分和保护河岸附近的缓冲区将有助于持续性地保护萤火虫种群,同时也助于生物多样性的保护(例如:其他无脊椎动物,植物,爬行动物,哺乳动物和鸟类。)
控制光污染
为了增加通过发光求爱的萤火虫种群的交配成功率,我们需要最小化其栖息地及其周围的ALAN。正在进行的研究,旨在研讨出具体的照明方案,包括光的颜色(波长)和亮度的调节,这将在一方面保障公共安全的前提下,也可提升萤火虫的繁殖率。但是,昆虫和其他动物的视觉敏感性不一而同,可能会限制调整的波长在特定群体所起的效果。与此相反,减少人造光的照射范围和持续时间,有益于一个极具文化特色和经济价值的夜间精灵。
减少杀虫剂的使用
在住宅花园,草坪和公园中,应尽量减少以美化为目的使用杀虫剂。而萤火虫幼虫期在废墟,地下或水下生活数月至数年,以至于它们常中招于杀虫剂。尽管只有极个别的研究查验了杀虫剂对萤火虫的直接影响,但常用的杀虫剂对多种非靶标生物(包括其他捕食性甲虫和幼虫萤火虫的猎物)具有不利影响。补充材料 补充的数据可于线上BIOSCI中可以查看
致谢
我们感谢所有受访者的意见,同时也感谢Larry Buschman,Ben Pfeiffer和Lynn Faust详尽分享了他们的见解,并感谢Sarah Hoyle提供了有关农药使用的信息。我们感谢Pedro Cardoso和David Wagner的详尽而周到的建议,极大地完善了此手稿。我们也十分感谢塔夫茨大学生物系的行政支持。
后记
昆虫数量下降及下降的因素近期得到了广泛的关注。萤火虫及发光生命是通过生物发光求爱行为的代表性昆虫,使之成为独特的文化符号,同时也被赋予了生态旅游特色的经济价值。
暂且不论萤火虫下降的迹象,我们对世界范围内约2000种萤火虫正不断减少的情况,全面核查其保护现状和所面临的威胁。我们对来自于不同的地理区域的专家进行了一个调查去分析对萤火虫种群和物种构成长久且最显著的预见性威胁。
栖息地消失,光污染,和农药使用被认为是最严峻的威胁,尽管各个地方大体上排名情况有所不同。我们的调查结果,伴随着对这些影响因子对于萤火虫种群的影响的全面综述。我们也讨论了这些风险因素有可能会降低特定物种应对特殊威胁时的抵抗能力。最后,我们倡议十分有必要建立一个长期密切跟踪濒危萤火虫的种群趋势监测计划。
流萤千千
青青流溪,点点萤火,汇聚成千千不息的银河,我们是一个旨在保护萤火虫及其栖息地的志愿团体,愿走过的路都伴随流萤千千。 目标:自下而上,保护萤火虫最后的栖息地。