
文丨木木科普官
编辑丨木木科普官
叶子暴露在强光下有时会导致光合效率降低,这种现象称为光抑制,植物对光抑制的敏感性取决于物种和生长光环境,遮荫植物低光生长植物比太阳植物,或高光植物更容易受到光抑制。
由于光抑制有可能降低生产力和植物生长,因此避免光抑制对于植物在自然栖息地的适应性和生存至关重要,照明时产生的有害活性氧参与了光抑制机制,单线态激发氧可以通过 相互作用生成与三重态激发的叶绿素形成于PSII反应中心。

超氧自由基不可避免地由梅勒反应,通过电子转移到O2在光系统 I. O 的突变2-导致形成过氧化氢和H2O2过渡金属离子产生羟基自由基,这是ROS中反应性最强的自由基。
在大麦中,叶子与一氧化碳一起孵育2-由于光合作用的光抑制,富氧空气呈黄色,这种现象显然与GL中观察到的现象相似。
植物材料
植物材料是在琉球大学校园内生长的榕树微果树L.f.金叶和榕树微果树。植物冠层上的光合成光子通量密度约为μmol m-2s-1,在此期间,在阳光充足的条件下,WT和GL在PPFD的生长条件上没有显著差异。
绿叶,在约100μmolm的光照条件下生长-2s-1最多,被收获并用作实验室实验作为遮荫叶,为了避免叶子的干旱,在整个测量过程中,叶柄连续保存在蒸馏水中,为了激活光合活性,将分离的叶子暴露在1000μmolm的照明下-2s-1实验前30分钟。
光照树叶,叶子被来自三个卤素块的白光照亮。在照明期间,用电风扇或恒温控制水浴将叶片温度保持在30°C,通过在光源前面放置金属丝网来获得各种光强度。
叶绿素荧光的测量
叶绿素荧光在大气条件下用PAM-2000叶绿素荧光计系统测量。在15分钟的暗适应期后,最小荧光 由微弱的红光确定,深色适应叶片的最大荧光在随后的白光饱和光脉冲。
最大荧光和照亮叶的稳态荧光在随后的白光饱和光脉冲上测量-2s-1,以确定 NPQ,PSII 和 qP 的产率,配备PAM-650的2000 nm发光二极管用于照明叶片作为光化光。
气体交换测量
一氧化碳2同化由CO的差异决定2同化室入口和出口之间的浓度,带有一氧化碳2气体分析仪的0.036% CO2和 21% O2,一氧化碳的测量同化速率在2300μmolm的照明下进行-2s-1,光响应曲线测量除外。
约 25 厘米2的叶子用于实验。快速搅拌腔室中的空气,以保持CO的高边界层电导率2扩散,O2使用气相沃克型氧电极系统,在 5% 一氧化碳中2和 21% O2 O的测量进化是在 4 厘米上进行的2在 600 μmol m 的照射下的叶盘-2s-1。
D1蛋白含量的测量
将叶(0.5g)在含有30mM磷酸钾,7mM苯基甲基磺酰氟和0%聚乙烯基聚吡咯烷酮的研磨介质中于5°C匀浆5秒。将匀浆与含有4%SDS的125ml 6mM Tris混合,然后在80°C下孵育3分钟以溶解蛋白质。
将样品冷却至室温后,通过以1,500 × g 离心30 s除去聚集体,将上清液与含有0.4M蔗糖,8M尿素,5mM EDTA和5%2-巯基乙醇的相同量的溶液混合。
使用含有6.0 M尿素的聚丙烯酰胺凝胶堆积和分离凝胶。在每个泳道中上样含有8 μg蛋白质的样品,使用针对D1蛋白产生的特异性多克隆抗体进行免疫印迹分析,蛋白质条带的强度使用NIH图像系统版本1.61测定。

现场光抑制
GL是热带无花果树榕树的栽培品种,原产于亚洲和大洋洲的亚热带/热带地区。GL的一个显著特征是黄叶含有大量的类黄酮,叶绿素和类胡萝卜素的含量可以忽略不计,黄叶中光合色素含量的减少可以通过从高光中遮蔽它们来增加。
因此,在遮荫条件下,GL表现出正常的绿叶,在形态上无法与WT叶区分开来。我们认为GL对光抑制的高敏感性可以解释这种现象。然而,没有直接证据表明GL对光抑制的敏感性。
光合性能
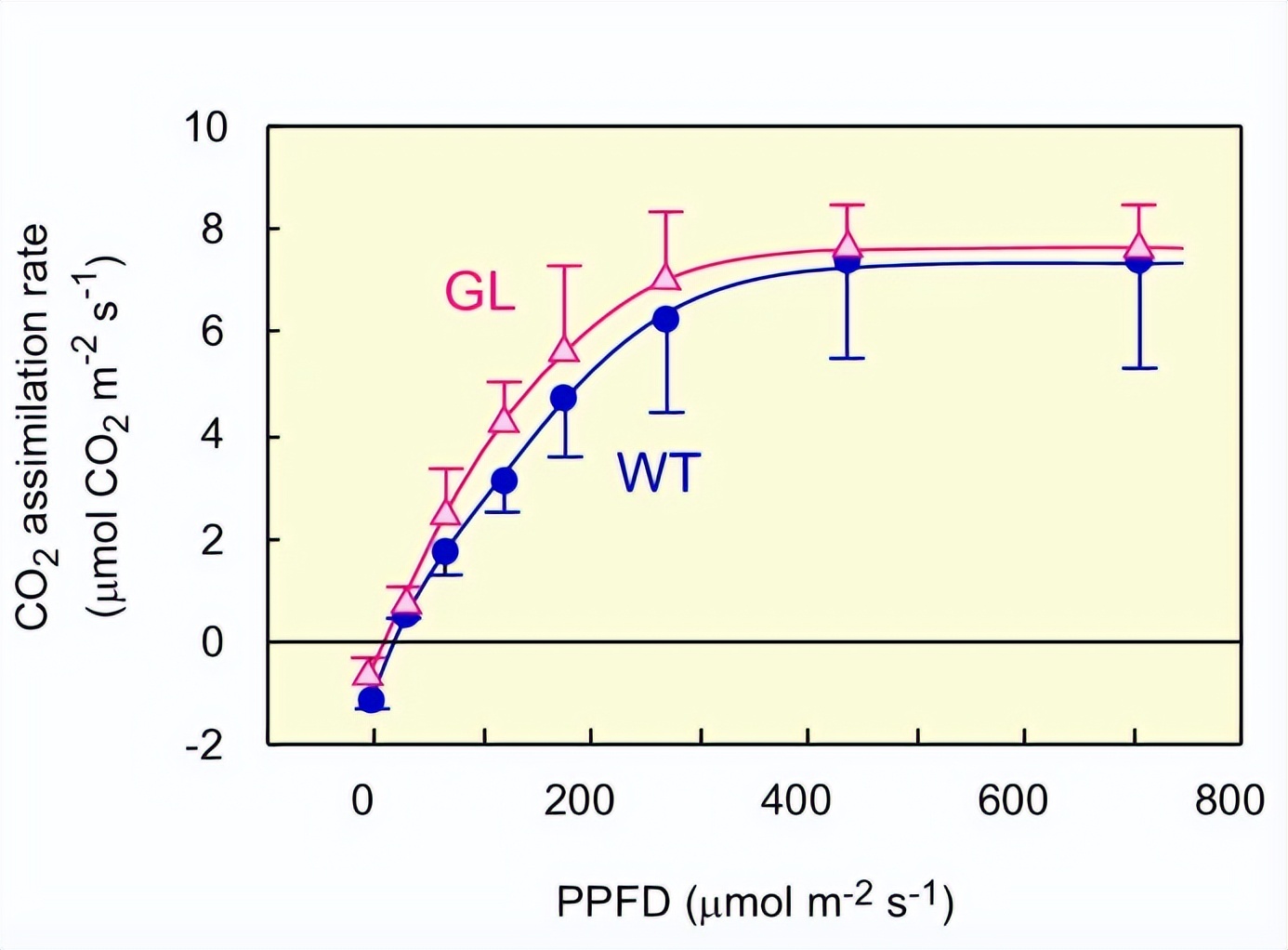
我们比较了GL和WT之间的光合性能,以确认GL的基本光合机制是正常的,GL和WT的遮阳叶都是绿色的,用于此目的。
GL遮荫叶叶绿素含量是0.58 g Chl m-2,在WT中为0.70克Chl m-2.GL显示出,与WT非常相似的光响应曲线,CO的最大活性没有实质性差异。这些结果清楚地表明,GL的光合能力与WT几乎相同,表明GL的光合机制在功能上没有缺陷。
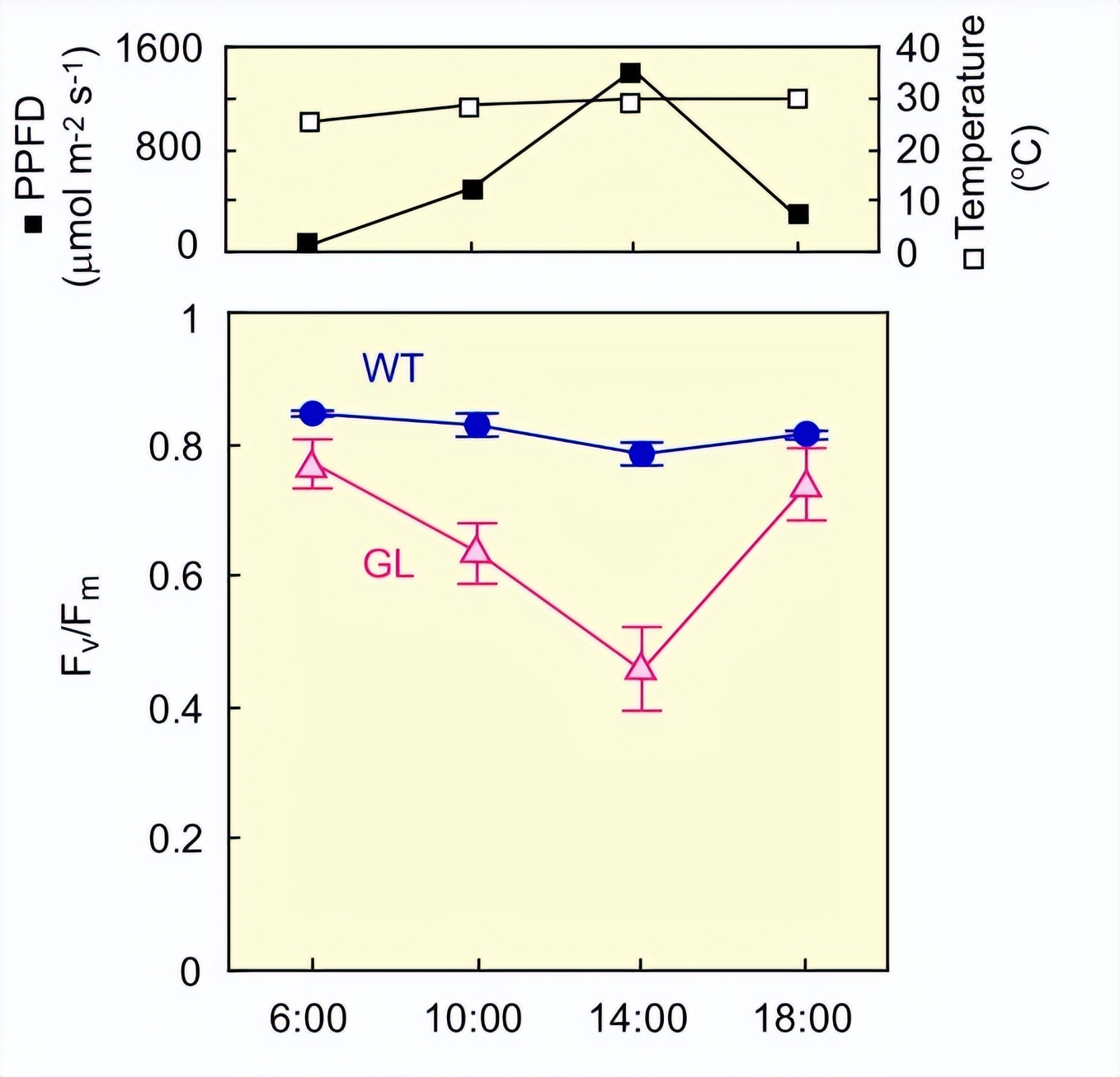
金叶遮荫叶的光抑制作用
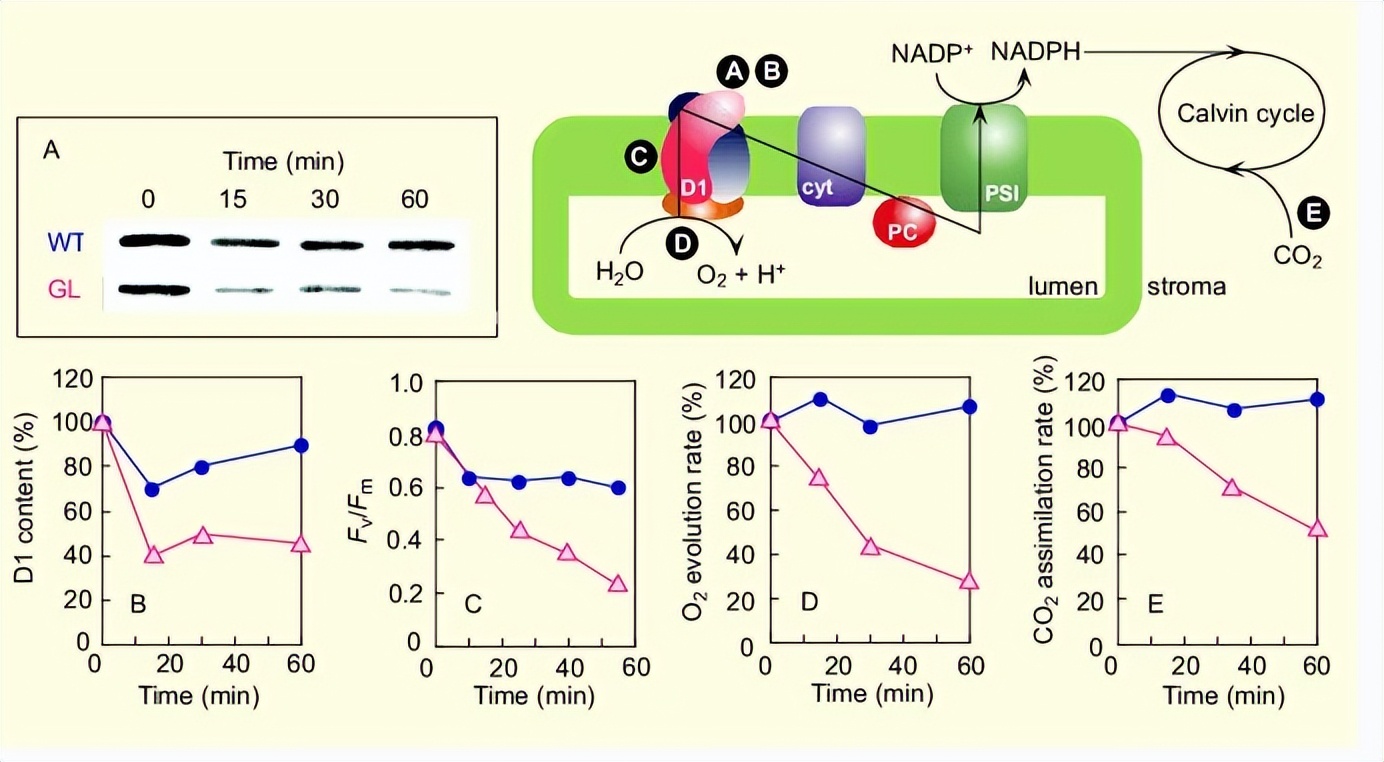
研究显示了高光对几个光抑制指标的影响:D1蛋白含量,Fv/Fm值,O2演化速率和一氧化碳2同化率。在高光照射之前,WT和GL遮荫叶在蛋白质基础上的D1蛋白质含量,没有实质性差异和 Fv/Fm值。
GL显示D1蛋白含量和F显著降低v/Fm高光照明时的值。与PSII活性的指标类似,净光合活性的指标在高光照明下也降低。
与在GL中观察到的这些反应相反,WT仅显示D1蛋白含量和F略有下降v/Fm值。演化与一氧化碳即使在高光照条件下,WT的同化率也没有变化。

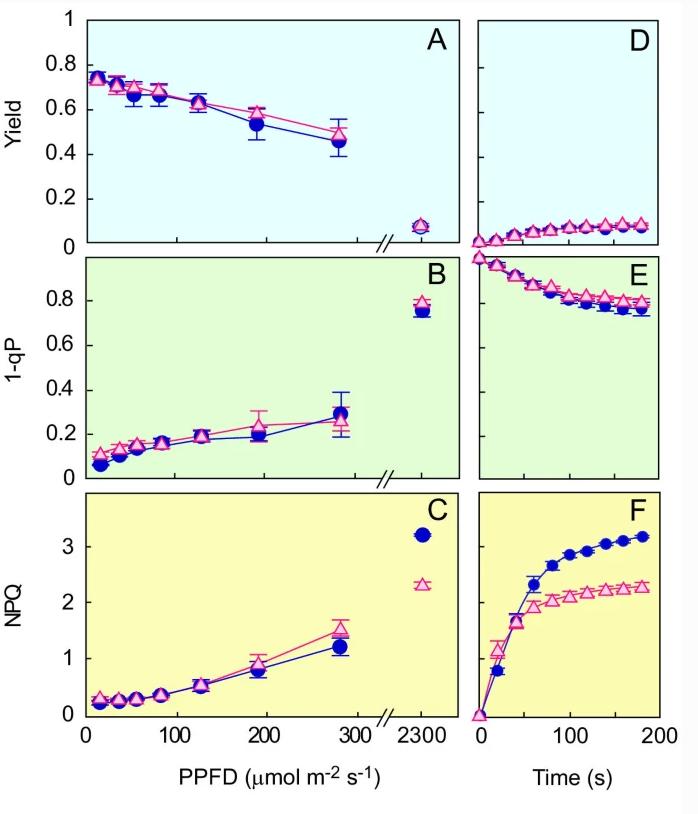
金叶NPQ容量降低
当植物暴露在超过光合作用能力的高光下时,多余的吸收光能可以安全地作为热量消散。这种能量耗散可以测量为PAM叶绿素的NPQ荧光分析,NPQ包括PSII的量子产率作为热能耗散的指标,PSII的光失活以及由于状态转变而导致PSII和PSI之间的光子接受度分布。
已知sate转变可以通过Q一个,可以测量为1-qP。GL和WT在产率和1-qP值方面没有显著差异。同时,电子传输速率的测量显示GL和WT之间的PSII光失活灵敏度基本上没有差异。
然而,与WT相比,GL显示出低得多的NPQ水平。这些结果表明,GL中的低NPQ可归因于散热机制的功能障碍。

研究表明,GL的光合活性在强光条件下极易受到光抑制。GL和WT之间的光合效率基本上没有差异,至少高达700μmol m-2s-1短时间,但GL中的NPQ低于WT中的NPQ,为2300 μmol m-2s-1。
拟南芥的npq1突变体显示出对光抑制的敏感性增加,但在短期光合O中不受影响演化与一氧化碳同化。对于NPQ的操作,已知PSII的光捕获复合物中的特殊叶黄素具有关键作用。
植物中NPQ的程度与位于类囊体膜管腔侧的紫黄质深度氧化酶由紫黄质形成的玉米黄质和蒽黄质水平密切相关。在光照过强的条件下,当类囊体腔的酸化达到临界阈值时,VDE被激活并通过蒽黄质将紫黄质转化为玉米黄质。
VDE的活性也受管腔中抗坏血酸浓度的调节,因为VDE只能利用抗坏血酸作为还原剂。已经表明,抗坏血酸可用性的降低严重影响体内VDE活性。由于类囊体膜没有任何抗坏血酸的主动转运机制,因此基质中抗坏血酸的浓度通过被动扩散机制决定了基质中的抗坏血酸。
在拟南芥中,已经清楚地表明,导致抗坏血酸缺乏的突变降低了体内NPQ性能。在野外条件下,GL的抗坏血酸含量在白天略有下降。因此,可以合理地假设抗坏血酸含量的降低是GL中NPQ功能障碍的原因。
抗氧化酶已经证明GL缺乏热稳定的DHAR活性,植物细胞具有几种表现出DHAR活性的酶:硫氧还蛋白,戊二糖还蛋白和二硫键异构酶。
从菠菜中分离出两种表现出DHAR活性的不同叶绿体蛋白,一种是库尼茨型胰蛋白酶*制剂抑**,另一种是特异性DHAR,由于两种叶绿体DHAR都比非叶绿体DHAR表现出,更高的热稳定活性,因此GL可能缺乏叶绿体DHAR。
在不含叶绿体的非光合器官中无法检测到热稳定活性,同时,我们预测叶绿体DHAR在抗坏血酸再生系统中起作用,以维持叶绿体中的抗坏血酸浓度,但DHAR在叶绿体中的生理意义仍存在争议。
由于基质MDAR的活性很高,因此在体内情况下可能不会发生从MDA到DHA和抗坏血酸的自发歧化。
因此,有一种观点认为,DHAR可能是抗坏血酸-谷胱甘肽循环中可有可无的,以保护叶绿体免受高光胁迫。然而,叶绿体中还有另一个位置,基质酶不能直接再生抗坏血酸,即腔腔空间。
在管腔中,叶黄素,α-甲胆酚和PSII氧化抗坏血酸导致DHA的产生,所有这些反应在强光条件下都可以促进。因此,基质DHAR对于减少管腔侧产生的DHA至关重要。
在这种情况下,叶绿塑料DHAR在生理上必须是光保护机制不可或缺的,特别是当叶子暴露在强光胁迫条件下时。这与先前的报道一致,即高光照条件下,拟南芥叶片中的DHAR活性增加。
因此,我们认为由于缺乏DHAR活性,GL的抗坏血酸可用性会因高光而降低,最终导致NPQ的性能降低。抗坏血酸不仅通过NPQ参与光保护,而且主要通过ROS清除参与光保护。
应该注意的是,GL对光抑制的高敏感性也可以解释为低ROS清除能力可能导致叶绿体中靶分子的光损伤和D1从头合成的抑制。
黄叶的产生机理仍然未知,GL的太阳叶含有少量的叶绿素和类胡萝卜素,但遮荫叶的叶绿素含量与WT相当,这意味着GL中黄叶的展示与长期高光照射有关,当GL的叶子暴露在强光下时,它们表现出明显的光抑制症状。
长期光抑制可导致叶绿素和类胡萝卜素等光合成色素的光漂白。NPQ中部分缺陷的npq1拟南芥突变体在强光条件下显示出叶绿素的光漂白,这种现象显然与GL中观察到的现象非常相似。

结论
与草本植物不同,叶子变黄现象不会导致衰老或细胞死亡。叶子中积累的类黄酮可以通过补充抗坏血酸 - 谷胱甘肽循环来保护组织免受氧化损伤, 除了ROS清除功能外,类黄酮长期以来一直被认为是有效的防晒颜料,特别是对紫外线辐射。
GL中增加的类黄酮色素可能起到ROS清除功能、紫外线防护和遮阳叶光衰减等作用。光抑制敏感性的机制,特别是GL叶片中的高耐光性仍有待充分阐明。
虽然很难将分子生物学技术应用于GL,但我们认为热带树木可以为研究在强光环境中生存的重要决定因素提供独特的机会。
参考文献
【1】Demming-Adams B,Adams WW:植物对高光胁迫的光保护和其他反应。植物生理植物分子生物学年鉴, 1992,43,599。
【2】Long SP,Humphries S,Falkowski PG:自然界中光合作用的光抑制。植物生理植物分子生物学年鉴, 1994,45,633。
【3】Powles SB:由可见光诱导的光合作用的光抑制。植物生理学杂志, 1984,35,15。
【4】Osmond CB:什么是光抑制?从遮荫植物和阳光植物的比较中获得的一些见解。在:光合作用的光抑制:从分子机制到场。编辑:Baker NR,Bowyer Jr. 1994,牛津,BIOS Scientific Publishers,1-24。
【5】Ball MC:低温下树木幼苗建立过程中光抑制的作用。在:光合作用的光抑制:从分子机制到场。编辑:Baker NR,Bowyer Jr. 1994,牛津,BIOS Scientific Publishers,365-376。