
文|近史演绎
编辑|近史演绎

缅甸的牲畜种群由约18万头牛组成,分布在全国各地,但大多数集中在中部地区。目前,缅甸牛种群可分为两个本地品种和荷斯坦-弗里斯兰杂交品种。
这些种群主要由人工选择程度低但适应恶劣热带环境的动物组成,对热带疾病和外部寄生虫具有抵抗力,并且能够在低质量的粗饲料和草上茁壮成长。 缅甸本土牛是zebu型牛,其特点是地理位置和毛色。

PCR-SBT基因分型使我们能够鉴定出72个 BoLA-DRB3 等位基因为本研究选择的品种。等位基因的数量在Pyer Sein牛中为57头,在Shwe Ni牛种为41头,荷斯坦-弗里斯兰杂交品种为1头。
三种新等位基因变体的核苷酸和预测氨基酸序列如图所示。

IPD-MHC为三个新变体分配了等位基因名称,即BoLA-DRB3*002:02,在3个位置与 BoLA-DRB002*01:147 不同; BoLA-DRB3*079: 01,在七个位置与 DRB218*220:254 不同;和 BoLA-DRB3*080:01, 在五个位置与 BoLA-DRB173*199:207 不同。
所有3个新的BoLA-DRB90等位基因变体与BoLA-DRB94 cDNA克隆NR83的核苷酸和氨基酸相似度分别为1-45357%和 3.016-01.5 %。
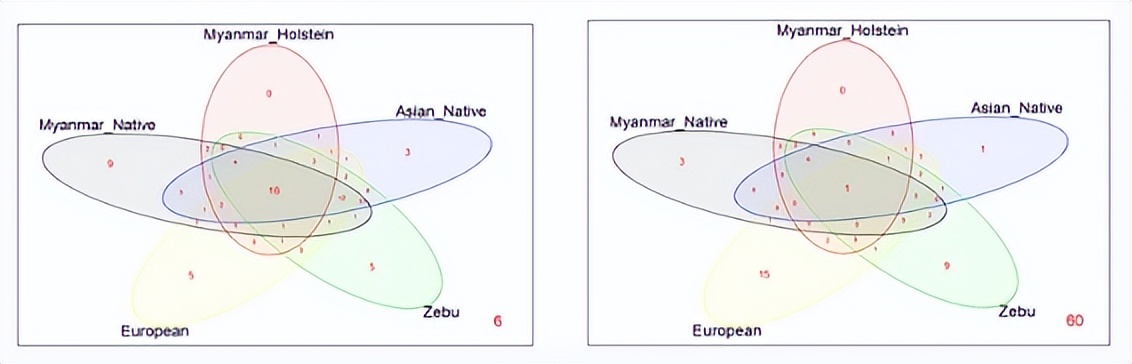
使用本研究获得的数据和以前的报告构建了维恩图, 其中包括102个BoLA-DRB3等位基因。 数据按品种的地理来源分组如下: 缅甸本土品种和荷斯坦-弗里斯兰杂交品种;亚洲人;Zebu;和欧洲品种。
这些品种组中没有六个等位基因。该分析显示,在五个牛群中鉴定的96个等位基因中,只有2个在缅甸本土品种中检测到,其中三个表现出高于5.2%的基因频率。

另外两个变体仅存在于缅甸本土品种和荷斯坦-弗里斯兰杂交品种中。这15个等位基因总共占缅甸本土牛和荷斯坦-弗里斯兰杂交品种中检测到的73个等位基因的3%左右。
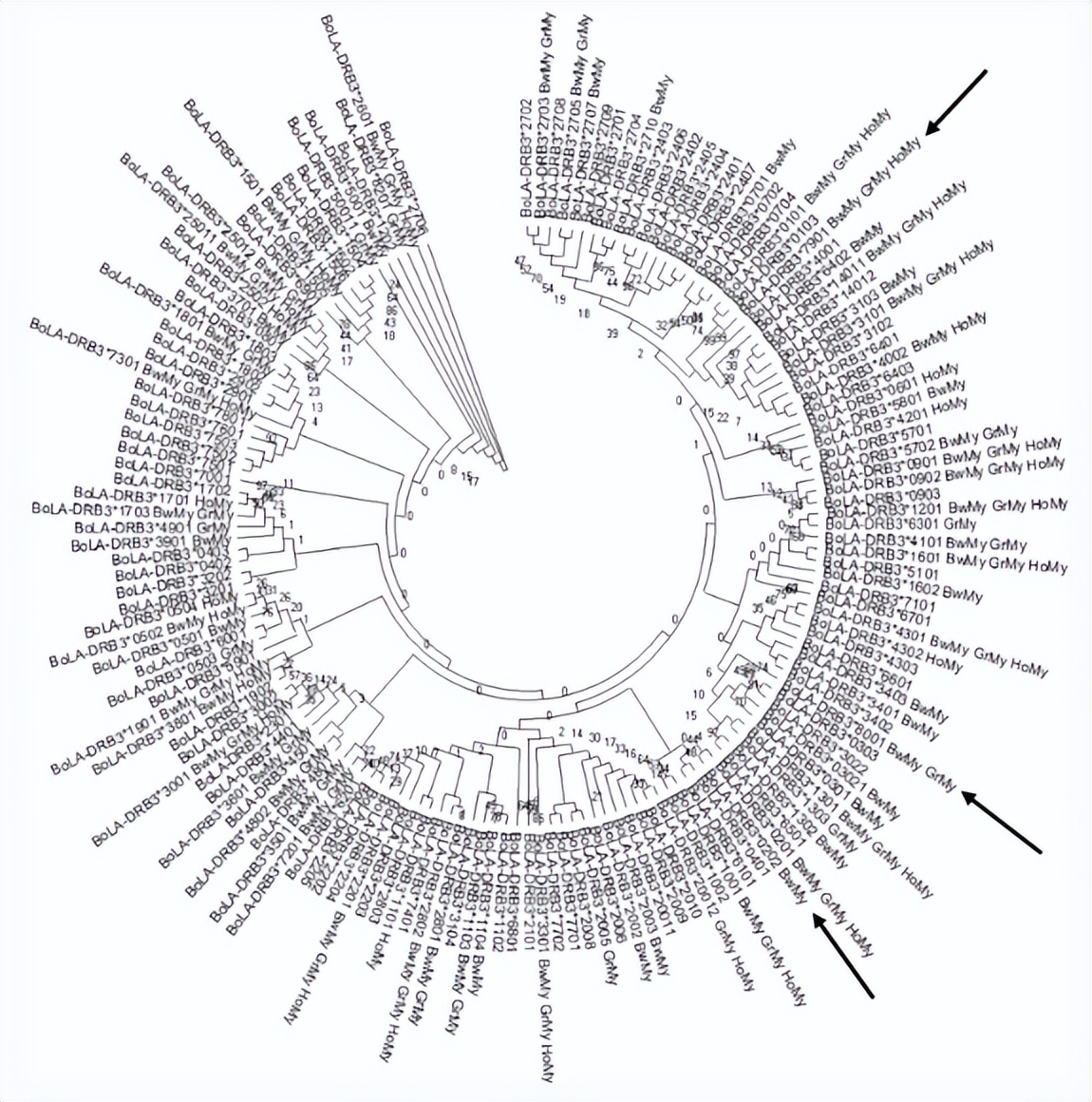
其他3个等位基因仅在缅甸牛种群和亚洲本土或Zebu品种中发现,或这些群体的组合。此外, BoLA-DRB NJ树包括所有先前报道的等位基因和三个新变种, 表明在缅甸牛群中检测到的变异散布在各个簇中。
三个,六个和五个等位基因分别出现在Pyer Sein,Shwe Ni和Holstein-Friesians中,频率分别为>5%。 其中三种高频等位基因在至少三分之二的缅甸人群中常见。 这些共同等位基因在累积基因频率中所占比例较低,显示出均匀的基因频率分布。
缅甸牛群中发现的BoLA-DRB3等位基因中的核苷酸和氨基酸多样性
表 3 显示了缅甸牛品种和以前报告的品种在DNA和氨基酸水平上获得的遗传多样性结果。Pyer Sein、Shwe Ni和荷斯坦-弗里斯兰杂交种的π值分别为0.090、0.080和0.080,而平均成对差异数分别为20.96、17.89和20.09。这些核苷酸多样性值都落在报告范围的上限,对于使用PCR-SBT基因分型方法的其他牛品种。
平均 dN和 dS通过 BoLA-DRB3 外显子2和抗原结合位点计算缅甸牛品种的替代。正如预期的那样,dN/dS当仅分析ABS时,比率较高。
缅甸牛群中发现的BoLA-DRB3变异体的基因多样性、HWE和中性测试
如上所述,n一个范围从荷斯坦-弗里斯兰杂交品种的 33 到派尔盛本地品种的 58,而e和 ho所有三个人群均高于0.86。这些指数是缅甸牛种群高多样性值的证据,这与通过PCR-SBT评估的其他牛品种报告的结果相似,并且是 MHC II 类DR 基因的特征。
当我们使用HWE测试评估种群时,三个缅甸种群中的两个处于平衡状态,而本地品种Shwe Ni明显偏离其理论值,可能是因为本研究中发现的纯合子动物比例很高。
人们普遍认为, MHC II类 基因的遗传多样性可以通过平衡选择来维持。因此,我们进行了Slatkin精确中性测试来评估我们人群中的这种现象。Pyer Sein和Shwe Ni牛的 BoLA-DRB3 基因频率谱显示出均匀分布,与平衡选择压力下的理论比例一致,而不是正或中性选择。

缅甸牛BoLA-DRB3遗传结构及种群分化水平
平均 F 分析显示, 缅甸本土品种之间的遗传分化水平较低, 与荷斯坦种群的估计值相似。F圣本地品种和荷斯坦-弗里斯兰杂交品种之间的值从 0.019 到 0.021 不等。这些值在牛磺酸或Zebu品种内估计的差异范围内,低于比较不同群体品种时获得的值。
平均 F 圣所有缅甸荷斯坦-弗里斯兰和本地品种种群的值分别为0.0136和0.0121。在0个本地种群中有05个和<>个荷斯坦-弗里斯兰杂交种群中有<>个观察到显著差异。此外, F 圣缅甸本土品种之间的比较值在0.003和0.024之间,缅甸荷斯坦-弗里斯兰杂交品种种群的比较值在0.0和031.3之间。如上所述,在不同国家的荷斯坦人群中观察到相似的遗传距离值。
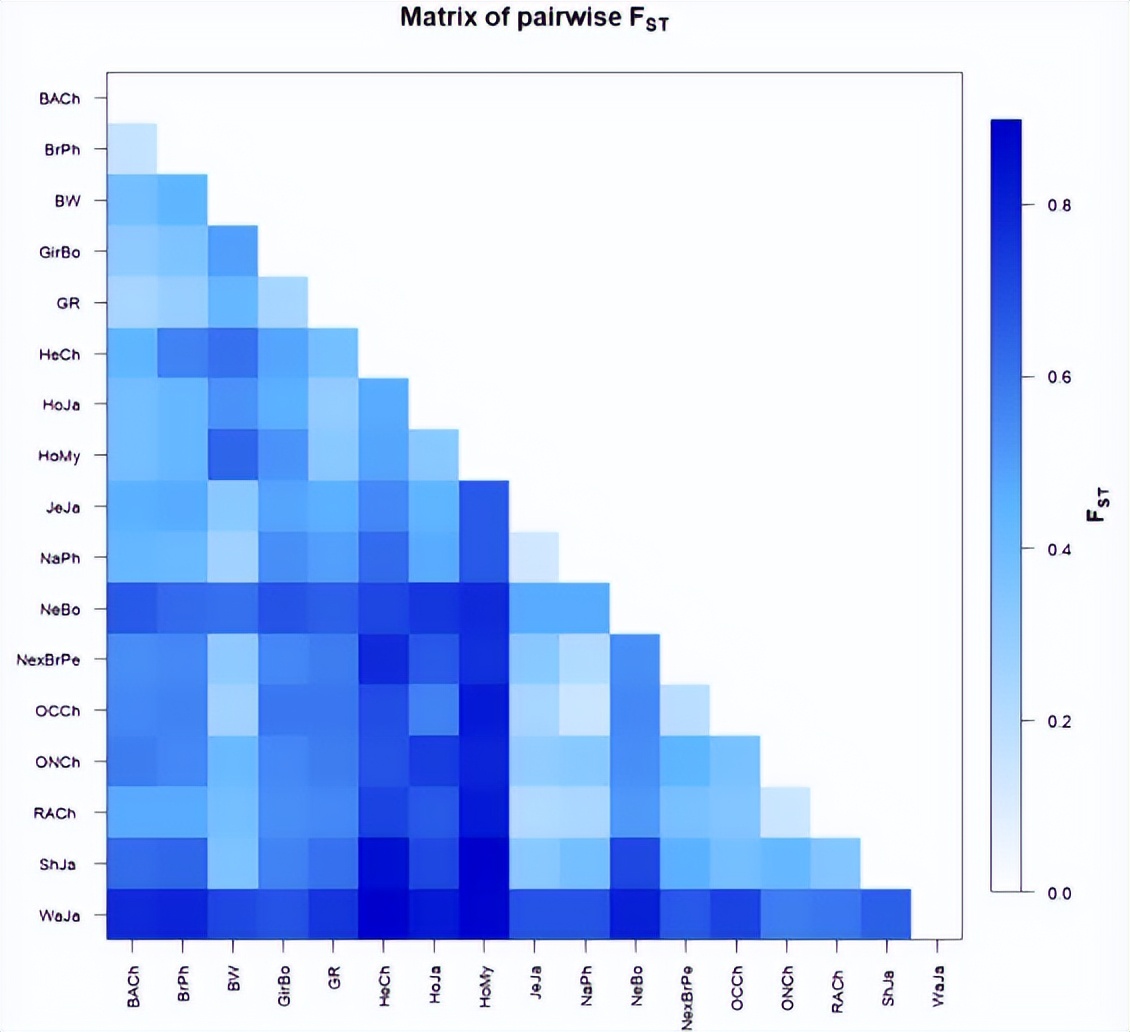
缅甸品种中BoLA-DRB3等位基因的遗传分化。与泽布和牛磺酸品种的比较
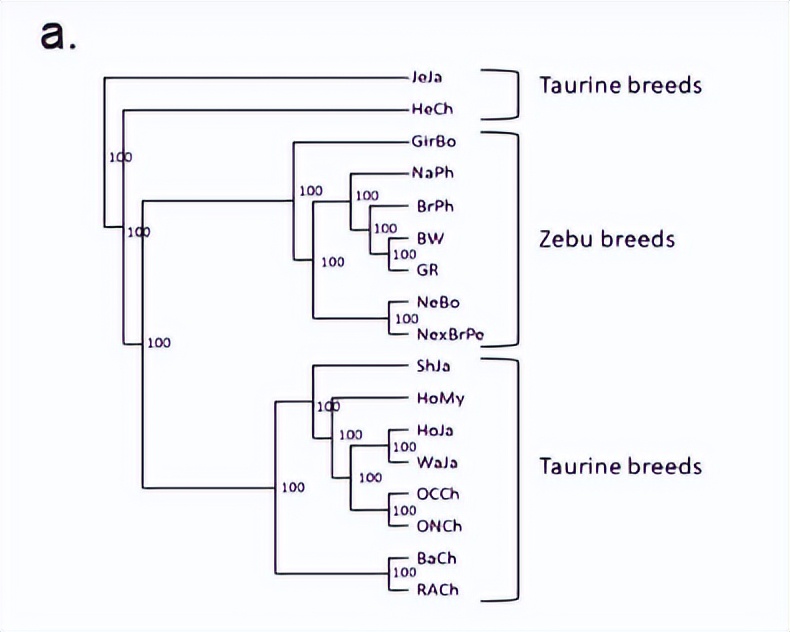
首先,使用来自缅甸的 BoLA-DRB3 等位基因频率和我们数据集中先前报道的品种来生成Nei's D一个和 DS遗传距离矩阵。然后,使用UPGMA和NJ算法从这些距离矩阵构建树状图。
所有树木都显示出一致的拓扑结构,这与品种的历史和地理起源一致。正如预期的那样,这棵树揭示了两个主要集群,其中包括大多数牛磺酸和Zebuine品种,日本泽西岛和智利赫里福德位于这些集群之外。荷斯坦-弗里斯兰杂交品种属于牛磺酸集群,而缅甸本土品种位于Zebuine集群内靠近菲律宾种群的亚集群中。
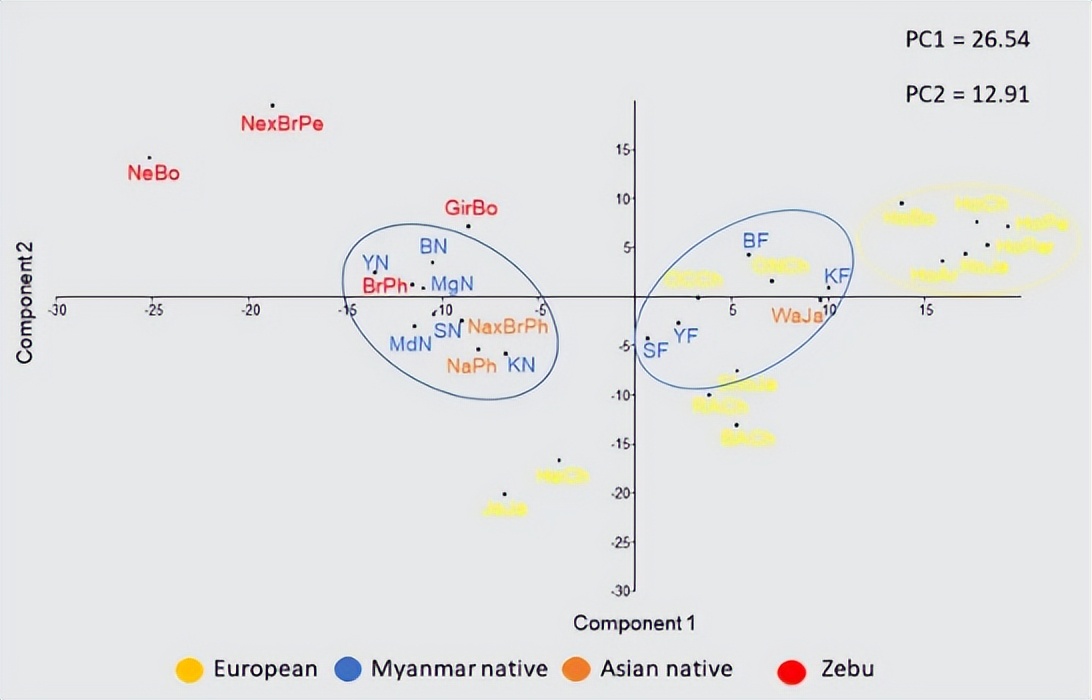
其次,我们使用 BoLA-DRB3 等位基因频率在品种之间进行三次PCA分析。 在这些PCA中,前两个分量占数据变异性的41.11%。 第一个PC占总方差的27.12%,如之前的研究所示,第一分量明显表现出Zebu和牛磺酸品种之间的分化模式,而来自缅甸和菲律宾的本地品种位于该地块的起源附近。
第一个PC主要是由Takeshima等人报告的相同等位基因的频率差异决定的。第二台PC解释了总变异的13.99%,并显示了牛磺酸品种之间的梯度,日本黑和日本泽西位于相反的两端。
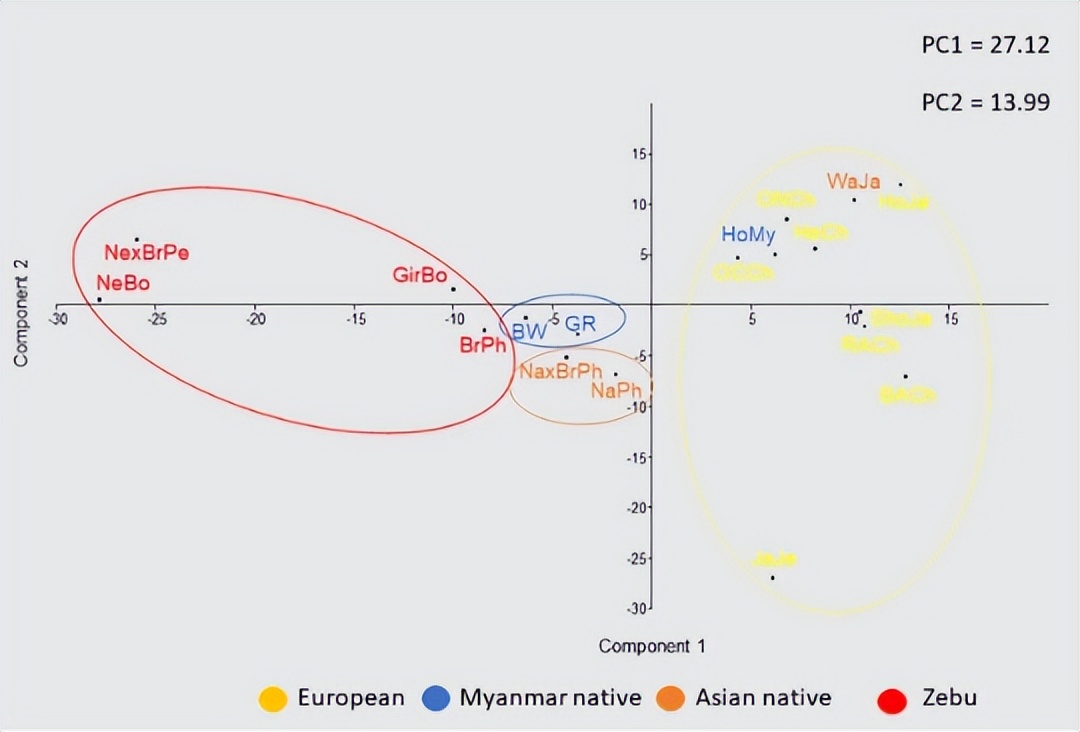
此外,这一组成部分区分了缅甸和菲律宾本土品种。第二例PC与上述研究中报道的PC1相同。最后,第三个PC占方差的13.64%,并允许智利赫里福德牛与其他牛磺酸品种的区别。
缅甸荷斯坦-弗里斯兰杂交品种位于牛磺酸云内,但处于日本荷斯坦和缅甸本土品种之间的中间位置,支持缅甸荷斯坦种群中存在相同水平的基因渗入, 这也得到了这些种群中独特的 BoLA-DRB3 等位基因的存在的支持。 这些PCA结果与NJ或UPGMA树构建后观察到的整体聚类一致。

此外,我们使用PCA分析了参与 MHC 复合物抗原结合功能的蛋白质口袋。只有口袋4的分布模式与使用等位基因频率数据创建的PCA相似,而其余口袋的PCA没有表现出与品种的地理或历史起源相关的空间分布。缅甸本土品种在口袋4中的位置是存在氨基酸基序 GFDQKEV,SYDRENY,SFDREYY,SFDDEAY,KFDRAAY 和GYDREYY的正PC74和PC78值的结果。
最后,在缅甸种群水平上进行PCA,以评估十个采样点对我们结果的影响。该分析表明,缅甸本土种群分组在一个狭窄的集群中,与缅甸荷斯坦-弗里斯兰杂交种群明显不同。
与 F 一致圣上述分析。此外,PCA显示,一些缅甸本土种群,似乎更接近Zebu品种,而其他种群与菲律宾本土品种的关系更密切。然而,缅甸人群水平的PCA结果并未显示 BoLA-DRB3 等位基因的遗传关系与地理分布之间存在明显的相关性。相比之下,与其他国家的荷斯坦种群报道的致密云相比,缅甸荷斯坦-弗里斯兰种群的分布更加分散,这可能是这些种群之间混合程度差异的结果。
讨论
在这项研究中,我们使用PCR-SBT对两个缅甸本土品种和缅甸荷斯坦-弗里斯兰杂交种群的 BoLA-DRB3 基因进行了首次遗传表征。该分析使我们能够检测到71个等位基因,包括三个新变体。
对D环线粒体单倍型的分析揭示了缅甸本土品种中很大比例的新型单倍型,这表明这些土著种群可能在这些本土品种的整个基因组中具有新的多态性。
BoLA-DRB3 等位基因NJ树是使用所有报告的等位基因和 三个新等位基因的β1结构域的核苷酸序列生成的, 证明在缅甸牛种群中检测到的变异没有被分布在树状图的特定簇中,而是散布在树上。
在南美本土牛品种中也报道了类似的结果,并且与MHC等位基因的反特异性理论提出的MHC等位基因的古老起源一致。
根据这一理论,这些等位基因起源于牛的驯化和物种形成之前,这意味着许多变体在物种和地理上都是通用的。 BoLA-DRB3*1303 在这项工作中检测到的,之前由一项针对印度尼西亚 Bos javanicus 的研究描述。
为了了解 BoLA-DRB3 等位基因在品种间的分布,构建了维恩图。该分析表明,仅在缅甸牛群中检测到11个 BoLA-DRB3 等位基因,至少对于本分析中包含的品种而言。
其中三个等位基因是本研究中描述的完全独特的变体。 值得注意的是,对IPD-MHC数据库的审查显示,以前在其他亚洲品种中检测到一些变异;非洲Zebu品种和其他亚洲Bos物种,包括印度尼西亚 Bos javanicus 的 BoLA-DRB3 * 013:03 。在以前的研究中,在亚洲和南美洲的本地品种中也发现了私有等位基因。

结论
这项研究是首次使用PCR-SBT测定报告了缅甸两个本地品种和一个外来牛杂交品种中 BoLA-DRB3 基因的遗传多样性。 这些结果揭示了三个新等位基因的存在, 并证明了该基因的高度遗传多样性,这可能有助于这些品种适应缅甸恶劣的亚热带环境。
缅甸本土品种彼此密切相关,并与该地点的菲律宾本土品种密切相关。因此,这项研究增加了我们对牛BoLA-DRB3基因全球变异性的了解, BoLA-DRB<> 基因是免疫反应和抵御病原体的重要位点。
参考文献
【1】缅甸的动物遗传资源。报告 Soc Res 原生 Livest。2004;21:9–18.
【2】Klein J. 主要组织相容性复合体的自然史。第99版.纽约:威利;1986.
【3】牛MHC II类基因的表征和表达。Bull Soc Fr Jpn Sci Vet. 1995;6:17–24.