
文 | 林夫
编辑 | 林夫

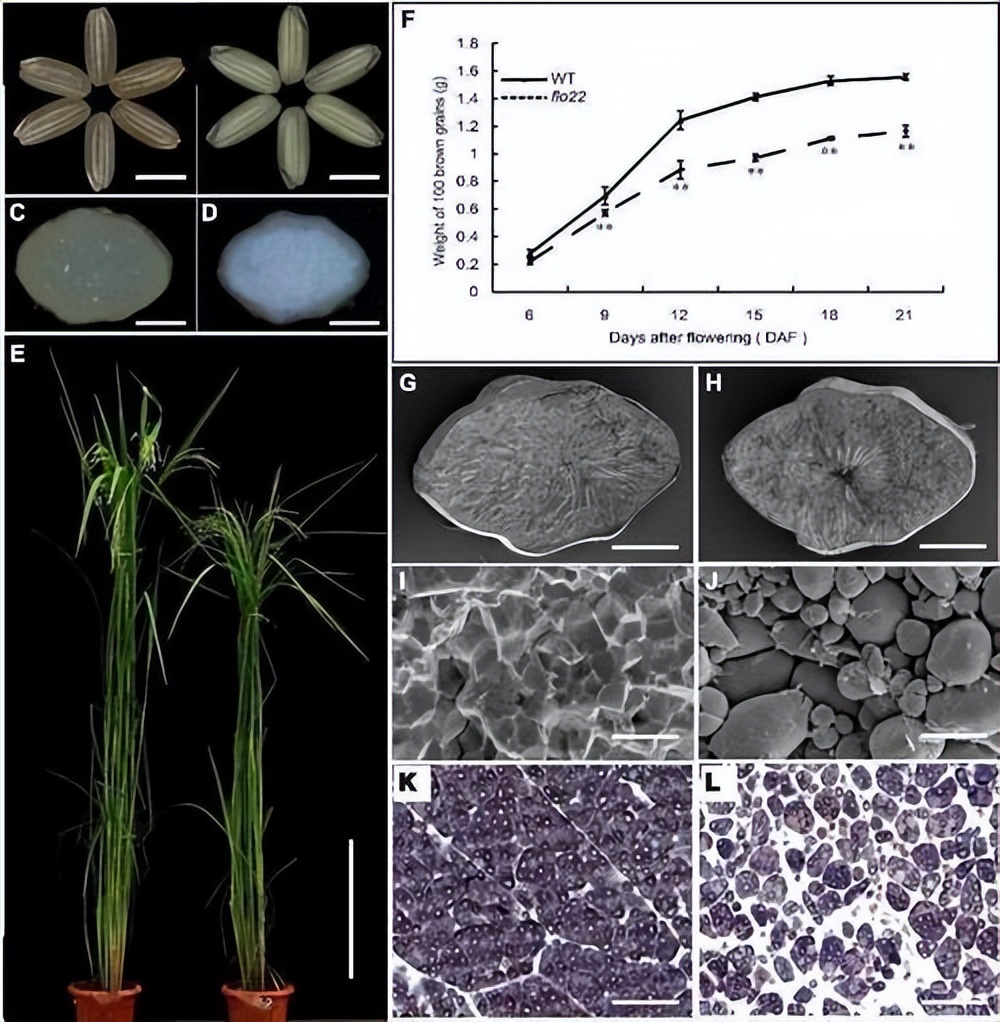
铜(Cu)是一种必需的微量营养素,在许多植物过程中起着关键作用,如呼吸、光合作用、氧化应激、氮和碳代谢、激素感知和细胞壁合成。

植物缺铜会导致生长迟缓、嫩叶发黄和发育缺陷,缺铜还会损害花粉的育性,降低生长速度、结实率和产量。

铜在人类健康中也起着重要作用。铜缺乏导致贫血和免疫缺陷。目前,全世界有20多亿人缺乏微量营养素,如铜、铁(Fe)和锌(Zn) 。

同样,过量的铜对植物也是有毒的,并引起植物根系生长抑制、生物量减少和植物坏死。铜毒性可使植物叶片变黄并降低光合速率。
铜毒性也会减少植物对铁的吸收,叶绿体中的类囊体膜也是铜毒性的主要目标。
铜离子的缺乏和过量对植物都是有害的,因此植物已经建立了精确的机制来平衡系统和细胞内铜离子的浓度。
Cu转运体在Cu2+的吸收、螯合、区隔化和代谢利用的关键步骤中起着重要作用。铜摄取相关蛋白主要包括铜转运蛋白家族,锌调节转运蛋白(ZIP)家族和黄色条纹状(YSL)蛋白家族。

目前,已鉴定的铜排泄蛋白主要来自p1b型ATP酶家族成员,即重金属ATP酶(HMAs)。
hma在植物金属运输中发挥作用。HMAs主要通过水解ATP输出金属阳离子。在植物生长过程中,HMAs不仅可以选择性地吸收和运输必需金属离子,如Zn2+和Cu2+,还可以选择性地吸收和运输非必需离子,如铅(Pb2+)和镉(Cd2+)。
当在酵母中表达时,CsHMA5.1和CsHMA5.2位于液泡膜中,并有助于酵母解毒Cu2+。
SvHMA5I和SvHMA5II分别定位于液泡膜和内质网,单独表达可提高拟南芥的铜耐受性。
Huang(2016)报道OsHMA4位于液泡膜中,可以将Cu储存在根细胞液泡中,减少水稻种子中Cu的积累。

拟南芥AtHMA5、水稻OsHMA5和OsHMA9位于质膜上,与细胞内铜的外排有关。
在本研究中,我们研究了OsHMA6的功能,发现OsHMA6可能是一种定位于质膜的Cu2+外排蛋白。

材料与方法
(一)水稻原料和生长条件
从水稻(Oryza sativa L. ssp)cDNA中扩增出OsHMA6开放阅读框(ORF)序列。粳稻的履历。
以武云粳7号水稻品种为材料,研究了OsHMA6和OsHMA9的表达模式及其对Cu和Cd的剂量响应。
采用南京农业大学试验站稻田不同生育期不同组织样品,研究了OsHMA6和OsHMA9的表达模式。

为了研究OsHMA6和OsHMA9表达对Cu和Cd的量响应,将14日龄幼苗在30℃/10 h黑暗(22:00-8:00)、22℃光周期和60%相对湿度的生长室内,在不同浓度的Cu(0.2、2、20、50、100、200和500 mol/L)或Cd(0、20、50、100和200 mol/L)下暴露3 d。
(二)RNA提取和实时PCR (qRT-PCR)
RNA的提取采用TRIzol试剂,用HiScript II Q Select RT SuperMix for qPCR (+gDNA wiper)试剂盒对处理过的总RNA进行逆转录(RT)。

使用2xT5 Fast qPCR Mix (SYBRGreenI)试剂盒进行三次定量分析,qRT-PCR引物见补充表1。
(三)OsHMA6在酵母中的异源表达
将空载体pYES2和构建载体OsHMA6-pYES2分别导入CM52酵母细胞,使用酵母转化试剂盒,根据制造商的协议,在合成葡萄糖培养基上选择转化子。
阳性克隆在SD-U液体培养基中培养至对数前期,浓缩后用无菌蒸馏水洗涤3次。
连续10倍稀释后,8个L细胞悬液分别在含有0、3和6 mmol/L CuSO4或0、10和20mol/L CdCl2的SD-U板上被检测到。在30℃下孵育3 d,然后评估生长表型。
用不同质粒转化CM52酵母菌株,在含有不同浓度Cu2+或Cd2+的SD-U液体培养基中进行生长测定。

制备过夜酵母细胞,用无菌蒸馏水调节600 nm处的光密度(OD)至0.5。然后,将20 L细胞悬液分别加入到每瓶含有0、3、6 mmol/L CuSO4或20mol/L CdCl2的20 mL SD-U液体培养基中。
在指定时间测定600 nm处OD值。在液体培养基中离心30 h收集菌株细胞,用无菌去离子水洗涤3次。
所有样品在80℃下干燥3 d,然后用电感耦合等离子体质谱(ICP-MS)测定金属浓度。
为了确定OsHMA6在酵母中的亚细胞定位,通过PCR将OsHMA6融合到YFP中,然后将融合片段插入pYES2载体中。
然后将获得的质粒转化为CM52酵母菌株,用共聚焦激光扫描显微镜观察YFP荧光。


结果
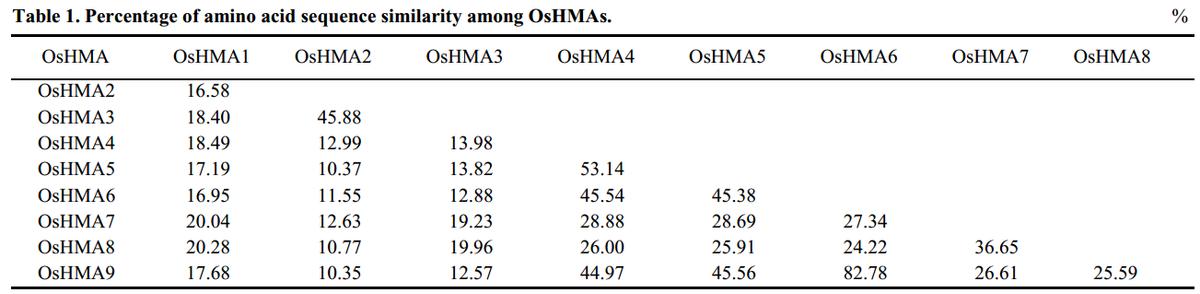
(一)OsHMA6和HMA家族成员的序列比对
为了研究HMA家族中OsHMA6基因的进化历史,我们对OsHMA和AtHMA家族进行了系统发育分析。
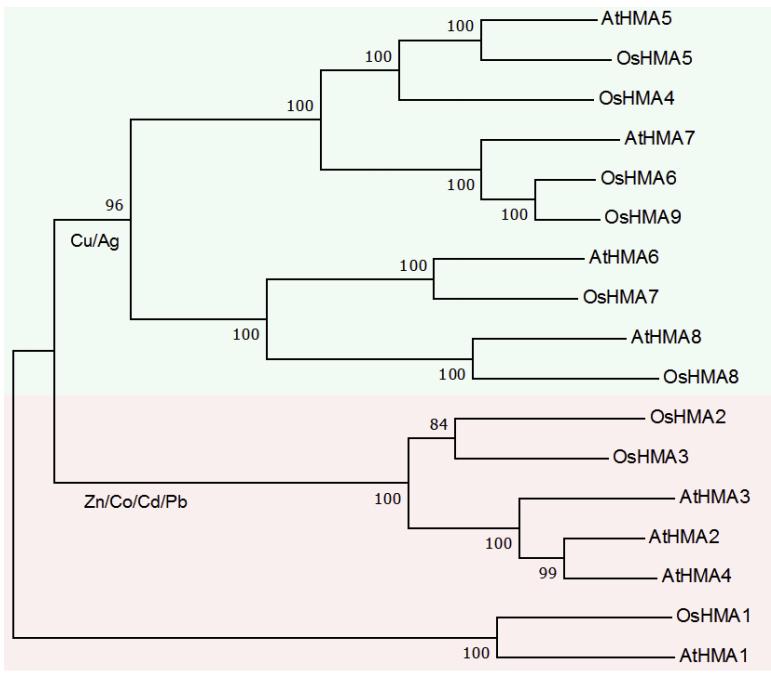
17个HMA基因被细分为两个分支,第一个进化支含有10个基因,属于Cu/Ag亚群,第二个进化支含有7个基因,属于Zn/Cd/Co/Pb亚群(图1)。
OsHMA6属于Cu/Ag亚群,与OsHMA9有着密切的进化关系(图1)。氨基酸序列比对结果如表1所示。
OsHMA6与OsHMA9的氨基酸序列相似性最高(82.78%)(表1),OsHMA6和OsHMA9具有相似的DNA结构,有9个外显子和8个内含子。
OsHMA6的CDS为3 039 bp,编码1 013个氨基酸;OsHMA9的CDS为3 012 bp,编码1 004个氨基酸。
OsHMA6蛋白的跨膜拓扑模型与OsHMA9相似。它们都有7个跨膜结构(补充图1-B)。
(二)OsHMA6和OsHMA9基因的表达模式
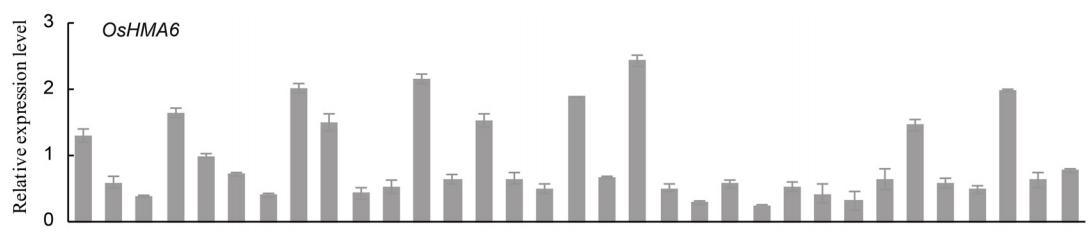
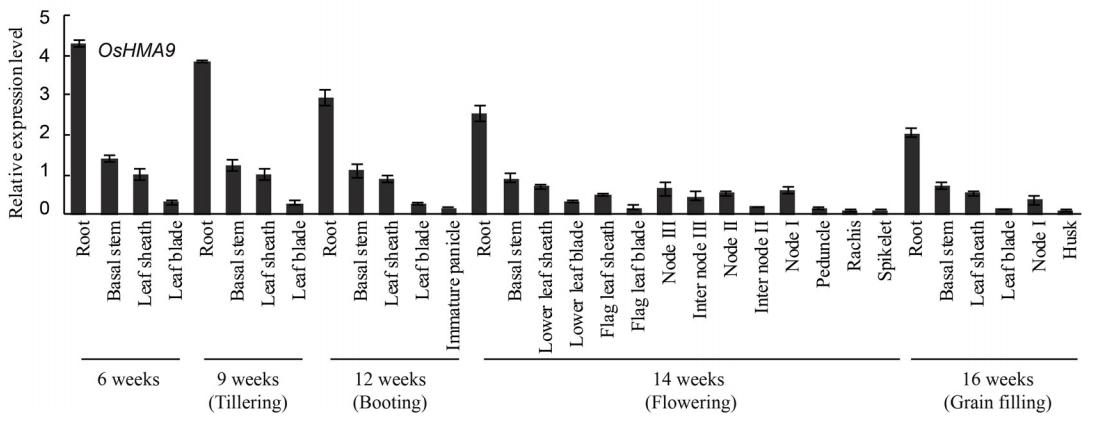
OsHMA9在根中表达最强烈,其次是基茎和叶鞘,在叶片中表达较少。在小穗中表达量非常低(图2)。
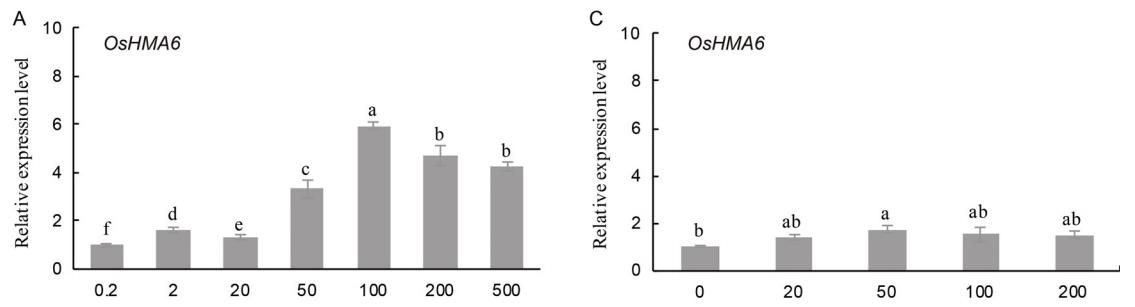
与对照(0.2 mol/L Cu2+)相比,高浓度Cu2+能显著诱导OsHMA6和OsHMA9的表达,其中OsHMA6在100 mol/L Cu2+时表达量最高,是对照的5.9倍(图3-A), OsHMA9在200 mol/L Cu2+时表达量最高,是对照的3.8倍(图3-B)。
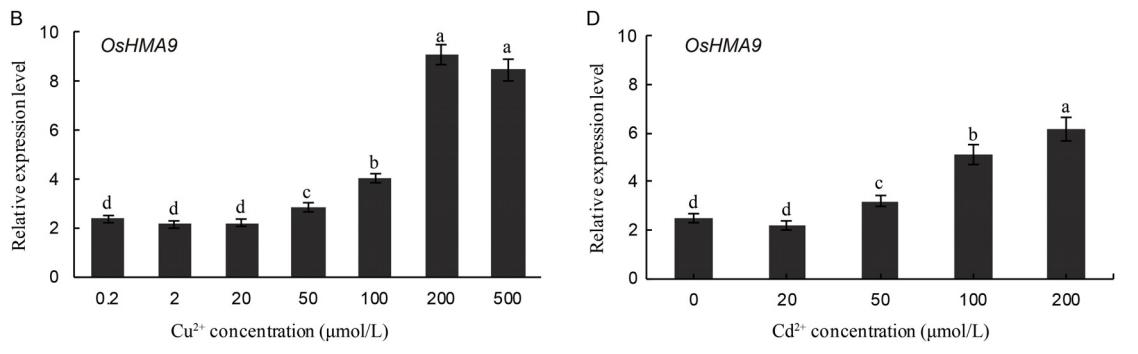
外源Cd2+的加入对OsHMA6的表达水平没有显著影响(图3-C),在200 mol/L Cd2+时,OsHMA9的表达量显著增加,是对照组的2.5倍(图3-D)。
(三)OsHMA6在酵母中的异源表达
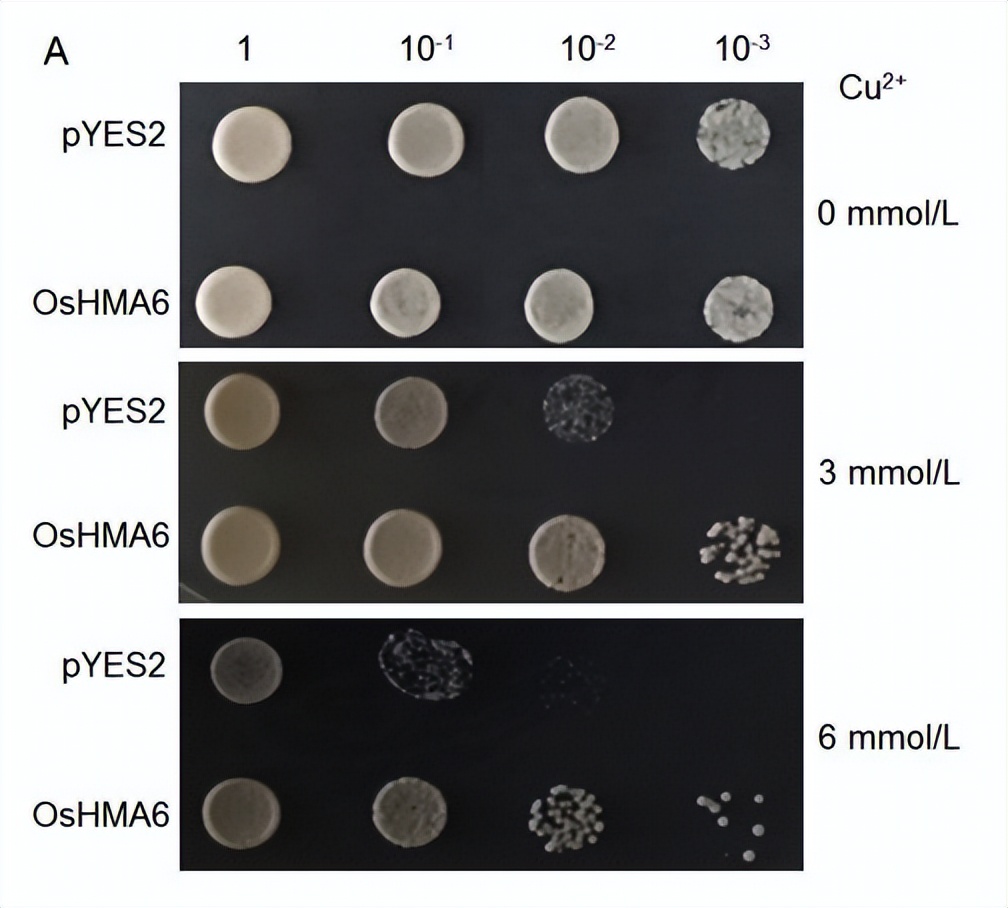
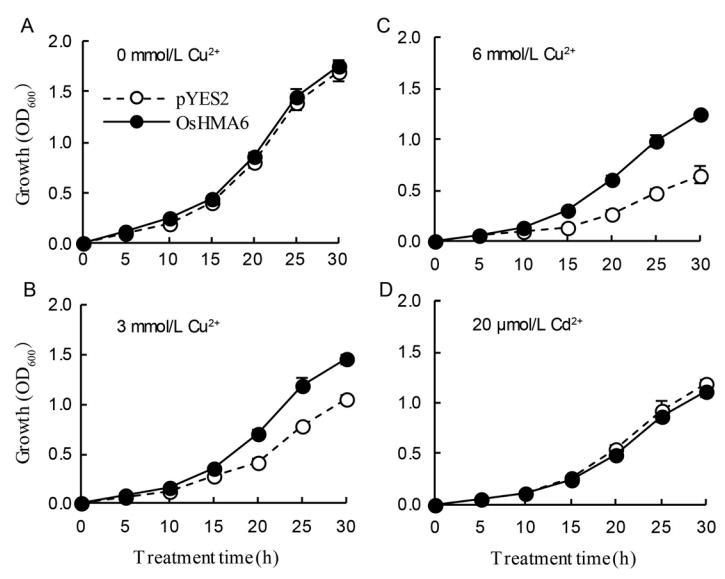
为了确定OsHMA6的Cu2+和Cd2+转运活性,我们将酵母菌株CM52中表达的OsHMA6与空载体pYES2进行比较。
结果表明,与空载体相比,OsHMA6在给药3或6 mmol/L Cu2+时能显著促进酵母菌株CM52的生长(图4-A),表明OsHMA6具有较高的Cu2+转运活性。
然而,在含有10或20 mol/L Cd2+的培养基上,OsHMA6和pYES2的生长没有显著差异(图4-B),这表明OsHMA6可能不具有Cd2+转运活性。
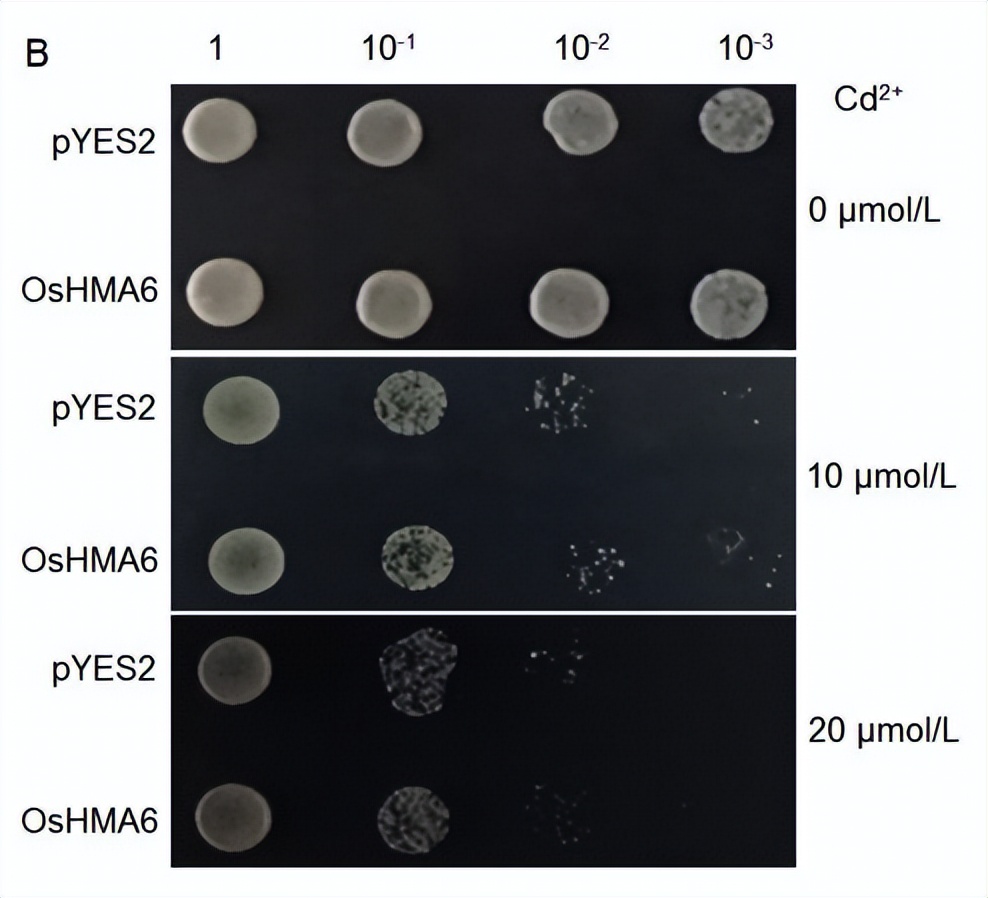
为了确定OsHMA6对酵母菌株CM52中Cu2+和Cd2+的敏感性,我们测量了转化后的CM52菌株在含有0、3、6 mmol/L Cu2+或20 mol/L Cd2+的SD-U液体培养基中的生长情况。
对照培养基和含有20 mol/L Cd2+的培养基中OsHMA6和pYES2的生长无显著差异(图5-A和-D)。
在含有3或6 mmol/L Cu2+的培养基中,OsHMA6在15h后的生长明显高于pYES2(图5-B和c)。为了进一步证实这一结果,分析了30小时应变中的金属浓度。
与pYES2相比,在3和6 mmol/L Cu2+处理下,OsHMA6中的Cu2+浓度分别下降了23.4%和30.3%(图5-F)。
然而,在20 mol/L Cd2+处理下,OsHMA6和pYES2的Cd2+浓度没有显著差异(图5-E)。
(四)OsHMA6蛋白定位于质膜
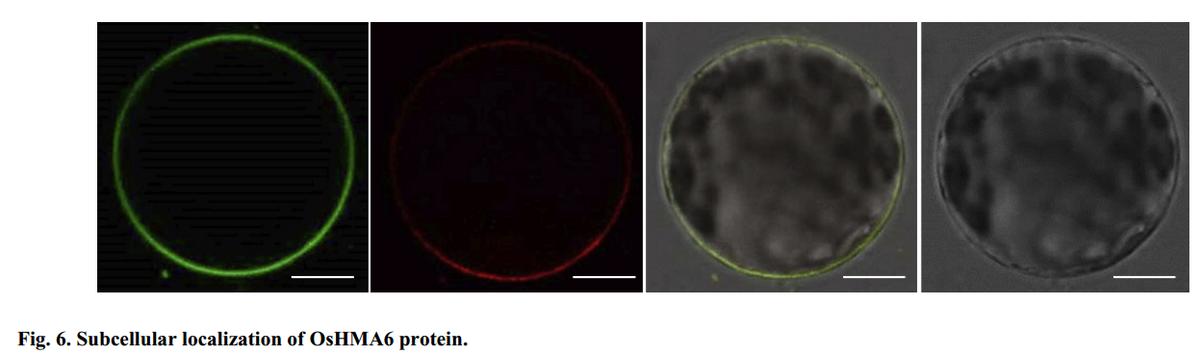
为了研究OsHMA6蛋白的亚细胞定位,构建了花菜花叶病毒35S启动子融合OsHMA6与GFP的载体,并将其转化到拟南芥原生质体中。
转化过夜后,观察原生质体中OsHMA6:GFP的表达情况。OsHMA6: GFP荧光与质膜染料FMTM4-64FX完全融合(图6)。
我们还将OsHMA6:YFP-pYES2转染到CM52酵母菌株中。
在半乳糖诱导表达后,我们观察到YFP荧光也定位在质膜上(补充图2)。因此,OsHMA6被鉴定为质膜定位蛋白。

讨论
铜是植物生长发育所必需的微量元素,铜必须通过根表皮和皮层的同心圆根细胞层,并被运输到中央柱。
植物需要将多余的铜储存在液泡中或运输到细胞外以确保其正常生长,并且运输到细胞外过程需要外排转运体。
因此,植物不仅需要铜转运体将铜吸收到细胞中,还需要一个外排转运体。稻田中铜的浓度从微摩尔到毫摩尔不等。
过量的铜供应会对植物产生毒性作用,OsHMA6和OsHMA9属于HMA家族中的Cu/Ag转运蛋白基团(图1),提示它们可能与Cu的转运有关。
OsHMA6和OsHMA9具有相似的内含子-外显子结构和蛋白质跨膜拓扑模型(补充图1)。OsHMA6和OsHMA9的氨基酸序列比对一致性为82.78%(表1)。
OsHMA9是一种位于质膜上的铜外排蛋白,敲除OsHMA9会影响水稻对高浓度铜的耐受性。我们推测OsHMA6可能是一种功能类似于OsHMA9的Cu外排蛋白。
与空载体pYES2相比,OsHMA6- pYES2在3或6 mmol/L Cu2+的固体培养基上能显著促进酵母菌株CM52的生长(图4-A)。
OsHMA6在含3或6 mmol/L Cu2+的液体培养基中培养15 h后,CM52的生长速度也明显快于pYES2(图5-B和c)。
这些结果表明OsHMA6具有较高的Cu2+转运活性。为了进一步证实这一结果,我们分析了菌株在液体介质中30 h的金属浓度。
与pYES2相比,在3 mmol/L和6 mmol/L Cu2+条件下,OsHMA6中的Cu2+浓度分别下降了23.4%和30.3%(图5-F)。
将OsHMA6:YFP- pyes2转染酵母株,YFP荧光定位在质膜上(补充图2)。
将OsHMA6:GFP融合载体转化到拟沙菌原生质体中,OsHMA6被鉴定为质膜定位蛋白(图6)。这些结果表明,OsHMA6可能是位于质膜上的Cu2+外排蛋白。
虽然OsHMA6和OsHMA9具有较高的氨基酸序列同一性和相似的Cu外排功能,但它们在水稻中的功能不同。

外源性Cd2+对OsHMA6的表达水平无显著影响(图3-C),而在200 ȝmol/L Cd2+时,OsHMA9的表达水平显著升高,是对照的2.5倍(图3-D)。
Lee(2007)报道在含有500 mol/L Cd2+的营养液中,OsHMA9敲除突变体的Cd浓度明显高于野生型,并且OsHMA9还具有Cd2+外排功能。
我们发现OsHMA6和pYES2酵母菌株在含有10或20 mol/L Cd2+的固体培养基上的生长没有显著差异(图4-B)和含有20 mol/L Cd2+的液体介质(图5-D和-E)。这些结果表明OsHMA6不具有Cd2+转运活性。
OsHMA6在不同生长阶段的各器官均有表达,包括小穗,其中叶片和根表达量最高(图2)。

OsHMA9在根中表达最强烈,在小穗中表达最弱(图2)。根系是吸收和运输重金属离子的主要器官,叶片是重金属离子的主要储存部位。
OsHMA9在木质部将Cu2+、Cd2+等根金属离子加载到木质部中起着重要作用。
AtHMA2和AtHMA4在小穗中的强表达表明,这两种转运蛋白在将锌转运到雄性生殖组织中起着特殊的作用。
拟南方植物中一种高亲和力的铜转运体COPT1在小穗中也有高表达,由于铜转运受阻,COPT1突变体在花粉发育中存在缺陷。
Athma5在花粉中表达同样强烈,并在拟南芥花药的金属运输中发挥作用。虽然这两个基因需要更详细的分析,但这些结果表明OsHMA6和OsHMA9在体内Cu2+平衡中发挥不同的作用。
参考文献
1.Abdel-Ghany S E, Müller-Moulé P, Niyogi K K, Pilon M, Shikanai T. 2005. 拟南芥叶绿体中的铜转运需要两个p型ATP酶. Plant Cell, 17(4): 1233–1251.
2.Alaoui-Sossé B, Genet P, Vinit-Dunand F, Toussaint M L, Epron D, Badot P M. 2004. 铜对黄瓜生长的影响及其与碳水化合物积累和离子含量变化的关系. Plant Sci, 166(5): 1213–1218.
3.Andrés-Colás N, Sancenon V, Rodriguez-Navarro S, Mayo S, Thiele D J, Ecker J R, Puig S, Penarrubia L. 2006. 拟南芥重金属p型ATP酶HMA5与金属伴侣相互作用,在根铜解毒中起作用. Plant J, 45(2): 225–236.
4.Andrés-Colás N, Perea-García A, Puig S, Peñarrubia L. 2010.铜运输的解除会影响拟南芥的发育,特别是在缺乏环境循环的情况下. Plant Physiol, 153(1): 170–184.
5.Argüello J M, Eren E, González-Guerrero M. 2007. 重金属转运p1b - ATP酶的结构与功能. BioMetals, 20: 233–248.