文 | 古梦今说
编辑 | 古梦今说
今天,小羊就来讲一下在核糖核酸的干扰下,病毒会对转基因的番茄植株有什么影响吧。
番茄黄叶卷曲病毒是一种单联 begomovirus(双生病毒科),是全球番茄生产严重减产的原因。
在阿曼,至少有五种不同的 begomoviruses 导致番茄疾病,包括 TYLCV。
不寻常的是,阿曼的 TYLCV 感染有时与 β 卫星(番茄卷叶 β 卫星;一种症状调节卫星)有关。
RNA干扰可用于在转录或转录后水平发展对 begomoviruses 的抗性。
产生了一种发夹 RNAi构建体,用于表达与番茄黄叶卷曲病毒-阿曼 的基因间区域、外壳蛋白基因、V2 基因和复制相关基因序列同源的双链 RNA。
最初,当用 TYLCV-OM 或 TYLCV-OM 和 ToLCB-OM 接种本氏*草烟**或番茄时,hpRNAi 构建体在病毒接种位点的瞬时表达显示可减少出现症状的植物数量。
用hpRNAi构建体转化 Solanum lycopersicum L. cv. Pusa Ruby,获得25个已确认的转基因系,并通过 农杆菌 介导的接种用TYLCV-OM和ToLCB-OM攻击。
对于除一个株系以外的所有株系,所有株系均无症状,接种 TYLCV-OM 导致一定比例 (≤25%) 的番茄株系出现感染症状。

对于 TYLCV-OM 和 ToLCB-OM 的接种,所有品系都显示一定比例的植物 (≤45%) 有症状。
然而,对于所有受感染的转基因植物,其症状较轻,植物中的病毒滴度低于受感染的非转基因番茄植物。
这些结果表明,RNAi 可用于在番茄中发展对双生病毒的抗性。
这种情况下的抵抗力不是免疫力,而是会降低感染的严重程度和病毒滴度。
此外,β卫星可能会损害抗性,增加最终表现出症状的植物的比例。
番茄是在热带和亚热带地区种植的第六大园艺作物。番茄黄叶卷曲病已成为许多地区番茄生产的关键限制因素。
自 Cohen在以色列首次报告 TYLCD 以来,引起该疾病的病毒已经传播,现在在世界范围内造成严重的产量损失。
番茄黄叶卷曲病毒 是一种单体Begomovirus,基因组为2.6-2.8 kb,由粉虱 Bemisia tabaci 传播。
双生病毒通过双链DNA 中间体复制,转录以双向方式从非编码基因间区域发生。

来自 OW 的单联 begomoviruses 的基因组编码六种蛋白质。病毒体意义中的基因编码外壳蛋白(CP:参与病毒在植物中和植物之间的移动)和前外壳蛋白(毒力决定因子,基因沉默抑制因子并参与植物中的病毒移动)。
互补义链编码复制相关蛋白(Rep:病毒 DNA 复制所需的唯一病毒编码蛋白,即滚环复制 起始蛋白)、转录激活蛋白(TrA:参与向上-调节晚期基因表达并可能是基因沉默的抑制因子)、复制增强蛋白和 C4 蛋白(可能作为基因沉默抑制因子的致病性决定因子)。
IR 区域包含病毒体链 DNA 复制的起点,该复制起点由预测的茎环结构组成,该结构包含保守的(在所有双生病毒中)非核苷酸序列 TAATATTAC和高亲和力Rep 识别序列称为 iterons 。
OW 中的 Begomoviruses 通常与一类称为 β 卫星的 DNA 卫星分子相关。
Betasatellites是真正的卫星,需要辅助病毒才能在植物中复制和移动以及在植物之间传播。
阿曼农业和渔业部于80年代末首次观察到TYLCV样症状,但直到2007年才将其病原体确定为TYLCV。
在阿曼鉴定的病毒对TYLCV表现出最高水平的核苷酸序列同一性(91%),并且是该物种的独特菌株;阿曼的TYLCV菌株。

此外,还鉴定出与TYLCV-OM相关的β卫星,其核苷酸序列同一性为88.5%,与从巴基斯坦分离的番茄叶卷曲β卫星有关。
在阿曼,农民使用三种主要的管理方法来控制 TYLCV;杀虫剂、物理屏障和部分抗性番茄品种。尽管采取了这些控制措施,但在阿曼的某些农场,TYLCV 的发病率有时可能达到 100% 。
常规育种被认为是获得抗性的最简单和最可靠的策略。然而,传统育种是一个耗时的过程,有时可能会由于连锁阻力而导致不良特性。
然而,常规育种在控制由白栗病毒或粉虱媒介引起的作物损失方面并不完全有效。
基因工程有可能解决这个问题,并为植物抗病毒提供了另一种途径。RNAi是一种进化上保守的机制,用于以序列特异性方式下调基因表达,由双链RNA 触发。
Begomoviruses 在转录水平(转录基因沉默)导致病毒 DNA 甲基化,在转录后水平(转录后基因沉默)导致病毒降解成绩单。
TGS 和 PTGS 都涉及一个 dsRNA 触发器,它被称为 Dicer 的 RNase 切割成短干扰RNA。

然后 siRNA 提供用于沉默的序列特异性。对于 PTGS,siRNA 被整合到酶复合物中,即 RNA 诱导的沉默复合物,它会降解与整合的 siRNA 同源的 mRNA。
以反向重复发夹结构的形式将与病毒同源的序列引入植物,是通过诱导基因沉默在植物中提供病毒抗性的有效方法。
基于 RNAi 的耐药性的成功依赖于沉默信号,该信号不仅限于单个细胞,而且可以从最初感染的细胞传播到更远的组织。
尽管双生病毒在其复制周期中没有 dsRNA 阶段,但它们确实会诱导病毒特异性 siRNA 的产生,并已显示可在受感染的植物中触发 PTGS。
感染木薯的双子病毒衍生siRNA在受感染的木薯中积累增加与疾病症状严重程度的相应降低有关,为RNAi作为植物双子病毒感染的适应性防御提供了线索。
与仅受 PTGS 影响的 RNA 病毒不同,双生病毒同时受 PTGS 和 TGS 影响。当产生与启动子区域同源的 siRNA 时,TGS 被触发,由于启动子序列的甲基化导致转录抑制。

TGS 在瞬时测定中显示出对绿豆黄花叶病毒有效,在该测定中显示受 MYMV 感染的黑革兰植物在接种靶向病毒启动子序列的 RNAi 构建体后完全从感染中恢复在红外中。
最近,基于RNAi的耐药性已成功应用于巴西的豆类中,以对抗 豆金花叶病毒 。转基因豆系在田间进行了测试,显示出对BGMV感染的免疫力。
这些转基因豆现在可供农民种植;作物对双生病毒的第一个商业化基于 RNAi 的抗性。
此处描述的研究调查了 hpRNAi 策略,作为通过靶向 TYLCV-OM 基因组的四个区域来控制阿曼 TYLCV 复合体的一种手段。
通过农杆菌介导的转化,将 hpRNAi 构建体转化到番茄植物中,讨论了多目标 hpRNAi 策略对 begomoviruses 产生抗性的潜力。
产生了包含 TYLCV-OM 的 Rep 基因、IR 区、V2 和重叠 CP 基因序列的多靶点 hpRNAi 构建体。
这包括 112 bp 的 IR(坐标 19-128)、161 bp 的 V2(坐标 129-288)、127 bp 的重叠 CP 和 175 bp(坐标 1760-1934)的 Rep。
575 bp 片段插入表达载体在正义和反义方向被一个内含子隔开形成一个发夹环结构。整个 RNAi 盒由双花椰菜花叶病毒35S 启动子表达。
用 TYLCV-OM 或 TYLCV-OM/ToLCB-OM 接种的本塞姆氏*草烟**植物在接种后 15 天显示出感染的最初症状,并且所有植物最终在 30 dpi 时出现症状。
总体而言,TYLCV-OM/ToLCB-OM 感染植物的症状比 TYLCV-OM 接种植物的症状更严重,叶片变小,叶片卷曲、变黄和叶脉肿胀更明显。
相比之下, 大多数本氏猪笼草 植物与hpRNAi构建体共同接种,TYLCV-OM或TYLCV-OM / ToLCB-OM保持无症状。
同样,更多的植物接种病毒和β卫星呈阳性,而不仅仅是病毒。PCR诊断表明,对于与hpRNAi构建体共同接种的植物,大多数植物都含有病毒。
但β卫星的存在增加了最终病毒阳性的植物数量;接种TYLCV-OM的植物16个中有30个,TYLCV-OM / ToLCB-OM接种的植物21个中有30个。

PCR阳性,无症状,hpRNAi构建体和TYLCV-OM或TYLCV-OM / ToLCB-OM接种植物的南方印迹。这表明没有杂交,表明这些植物中的病毒和β卫星水平低于杂交检测的阈值。
在不存在或存在ToLCB-OM的情况下,所有与hpRNAi构建体和TYLCV-OM共同接种的番茄植株均无症状。这些植物继续正常生长并开花。
相比之下,接种TYLCV-OM时,非转基因番茄植株出现叶面黄变和轻度卷曲30 dpi,而接种TYLCV-OM / ToLCB-OM的植物发生更严重的叶面黄变,发育迟缓和30 dpi的叶面卷曲。
番茄植株的南方印迹分析检测到接种TYLCV-OM和TYLCV-OM/ToLCB-OM的植物双子病毒复制的典型病毒DNA形式。
探测β卫星存在的印迹显示ssDNA杂交用于接种TYLCV-OM / ToLCB-OM的番茄。相比之下,在hpRNAi构建体存在的情况下,接种TYLCV-OM或TYLCV-OM / ToLCB-OM的番茄植株均未与检测到的病毒或β卫星探针杂交。

除单一植物外,在与hpRNAi构建体共接种的番茄植株中,未通过PCR检测到TYLCV-OM和ToLCB-OM,而PCR反应中产生了指示病毒/β卫星的特定DNA条带,其中包含从接种了TYLCV-OM或TYLCV-OM / ToLCB-OM但没有hpRNAi构建体的有症状植物中提取的DNA。
石蒜链球菌 L.cv 红宝石植物通过 农杆菌 介导的转化与hpRNAi构建体进行转化。共获得11株卡那霉素耐药初级转化番茄植株。
然而,使用引物CS-For/CS-Rev进行PCR分析,指导查耳酮合酶内含子约1100 bp片段的扩增,表明只有9种植物含有转基因。
这9家工厂进展到T1通过自花授粉产生,用于评估对TYLCV-OM和TYLCV-OM/ToLCB-OM的抗性。所有推定的转基因品系(每个品系20个种子)在500 mg/L卡那霉素选择培养基上发芽,然后转移到盆栽土壤中。
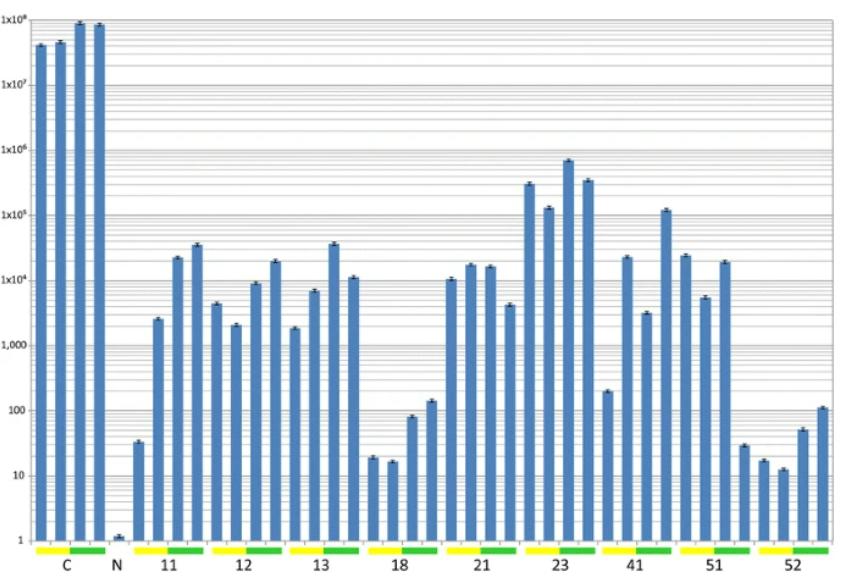
大多数接种的转基因植物仍然没有症状。
除了第41系的植物,没有接种TYLCV-OM的植物出现症状外,所有品系在接种TYLCV-OM后显示高达25%的植物出现轻度症状,高达45%的植物接种TYLCV-OM / ToLCB-OM为60dpi这里介绍的结果表明。

这与已知的β卫星一致,在大多数情况下,β卫星会增加植物中的辅助病毒DNA水平。β卫星诱导的病毒DNA水平增加的确切原因尚不清楚。
对接种番茄植株的南方印迹杂交分析在有症状的非转基因植株中检测到高水平的病毒和β卫星。然而,除了一种检测到低水平β卫星的植物外,在接种的转基因植物中没有检测到病毒或β卫星DNA。
总之,在核糖核酸的干扰下,会降低感染的严重程度和病毒滴度。