在阅读此文前,诚邀您请点点右上方的“关注”,既方便您进行讨论与分享,还能及时阅读最新内容,感谢您的支持。
Hopwood曾经报道天蓝色链霉菌A3(2)菌株中决定次甲基霉素生物合成和对该抗生素抗性的SCP1质粒能通过接合作用转移到变青链霉菌(S.lividans)及小小链霉菌(S.parvulus)中,使它们获得合成次甲基霉素和抗该抗生素的能力,这是迄今为止我们所知道的与抗生素生物合成有关并具有种间接合转移能力的第一个链霉菌质粒。
在其它已知的链霉菌质粒中是否也具有类似于SCP1的特性?这是一个使我们感兴趣的问题。
我们以前的工作,从遗传上证明了庆丰链霉菌中存在质粒SQP1,它参与庆丰霉素(以后简称Qm)的生物合成并和该菌株对Qm的抗性有关。
这个质粒能以很高的频率通过种内的接合作用从SQP1+菌株转移到SQP1-菌株,使SQP1-受体菌重新成为能产生Qm并抗Qm的转移接合子。
那末SQP1质粒能否在链霉菌的不同菌种间进行转移?它对Qm生物合成遗传控制的情况又是怎样?我们以庆丰链霉菌SQP1+的菌株作为供体和其它不产生Qm的链霉菌进行种间接合转移试验,检查配对的受体菌所发生的表型变化,以判断SQP1质粒是否能进行种间转移,并分析SQP1质粒在Qm生物合成途径中可能起的作用。
本文报道了SQP1质粒在庆丰链霉菌A201菌株和井冈链霉菌VA4菌株间的种间转移及接合子P2菌株的一些特性。
材料和方法
(一)菌株
庆丰链霉菌(Sireptomyces qingfengmyceticus) A201[Ilv-QmrSQP1+]菌株,由原养型M15菌株经亚硝基胍诱变获得[3]的异亮氨酸、缬氨酸缺陷菌株。
井冈链霉菌(Streptomyces hygroscopicus var jinggangensis) VA4[His-Qms]菌株,由上海农药研究所提供的原养型75号菌株经NTG诱变获得。
指示细菌:枯草杆菌AS 1.140。
(二)培养基
庆丰链霉菌斜面孢子培养基(CM),最低合成培养基(GA),Qm效价测定培养基等均见前文报道。
井冈链霉菌斜面孢子培养基,发酵培养基均参照上海农药研究所的报道。
(三)接合转移试验
配对二亲株经分离纯化分别接种在各自斜面上,VA4置37℃,A201置28℃培养4一6天,然后将新鲜孢子混合接种在同一支CM斜面上,28℃培养6天,待孢子成熟,用无菌生理盐水(含0.1%的吐温80)洗下混合孢子,适当稀释后涂布于补充组氨酸(70γ/ml)的最低培养基平板上,挑取VA4单菌落,用琼脂块法鉴定各单菌落抑制枯草杆菌的能力。
(四)井冈霉素(以后简称Jm)硫酸苯酚比色测定法参照上海农药研究所的报道。
(五)抗菌物质的分离
试验菌株的新鲜孢子接种于井冈霉素发酵培养基中,于37℃转床(240转/分)培养四天,发酵液用草酸调pH至4左右,过滤除去菌丝体,滤液通过强酸型阳离子树脂Dowex50 (Na+)柱进行交换,用2N HCl洗脱,分部收集(每管3ml),逐管测定硫酸苯酚显色反应及274毫微米波长处的光密度。
取硫酸苯酚试剂呈阳性反应的流段(称为P2A)及波长274毫微米紫外吸收峰位的流段(称为P2B),分别以阴离子树脂711 (OH-)中和,减压浓缩,4℃冰箱贮存,供试验用。
(一)配对链霉菌的选择
庆丰链霉菌SQP1+菌株因其能产生Qm而对许多种微生物如革兰氏阳性、阴性细菌、红酵母、大肠杆菌等产生抑制作用。
考虑到实验时便于从混合培养的受体菌孢子中利用对细菌的抑制作用来鉴别获得SQP1质粒的接合子,我们选择具备下列特性的链霉菌作为种间配对的受体菌:(1)不产生有抑制细菌作用的抗生素。
(2)对庆丰霉素敏感。
(3)具有与庆丰链霉菌不同的外观形态特征。
对一些链霉菌进行了试验后,认为井冈霉素产生菌符合这些要求,它和庆丰链霉菌特征的比较,。

表1指出VA4不产生能抑制细菌的物质,同时对Qm敏感,这些特性相当于SQP1-的菌株,因之在我们的整个实验中选择井冈链霉菌的组氨酸缺陷衍生株VA4作为SQP1质粒种间转移的受体菌。
图1 配对亲株和接合子的菌落形态培养基为GA培养基补充有氨基酸(a.组氨酸、
(二)SQP1质粒的种间转移
按方法中所述的实验步骤将A201[Ilv-His+SQP1+]菌株和VA4[Ilv+His-SQP1-]菌株在CM培养基上进行混合培养,并在含组氨酸的最低培养基上检出混合孢子中的受体菌VA4,用琼脂块法鉴定对枯草杆菌的抑菌活性,经过多次实验表明有2.22—8.86%的VA4菌落获得了抑制枯草杆菌的能力,并同时表现对Qm的抗性,接合子除了获得亲株VA4所没有的抑细菌能力外,其余特性如菌落的外观,分泌井冈霉素,对水稻纹枯病菌产生异常分枝等均和亲株VA4完全相同。
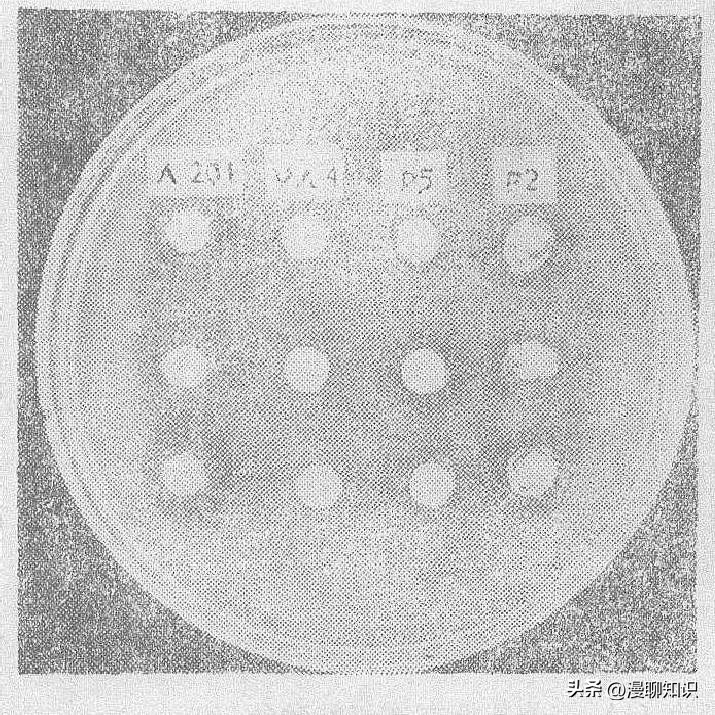
图2 配对亲株和接合子对枯草杆菌的抑制(琼脂块法)
图3亲株VA4和接合子对水稻纹枯病菌的抑制
(三)接合子的遗传稳定性
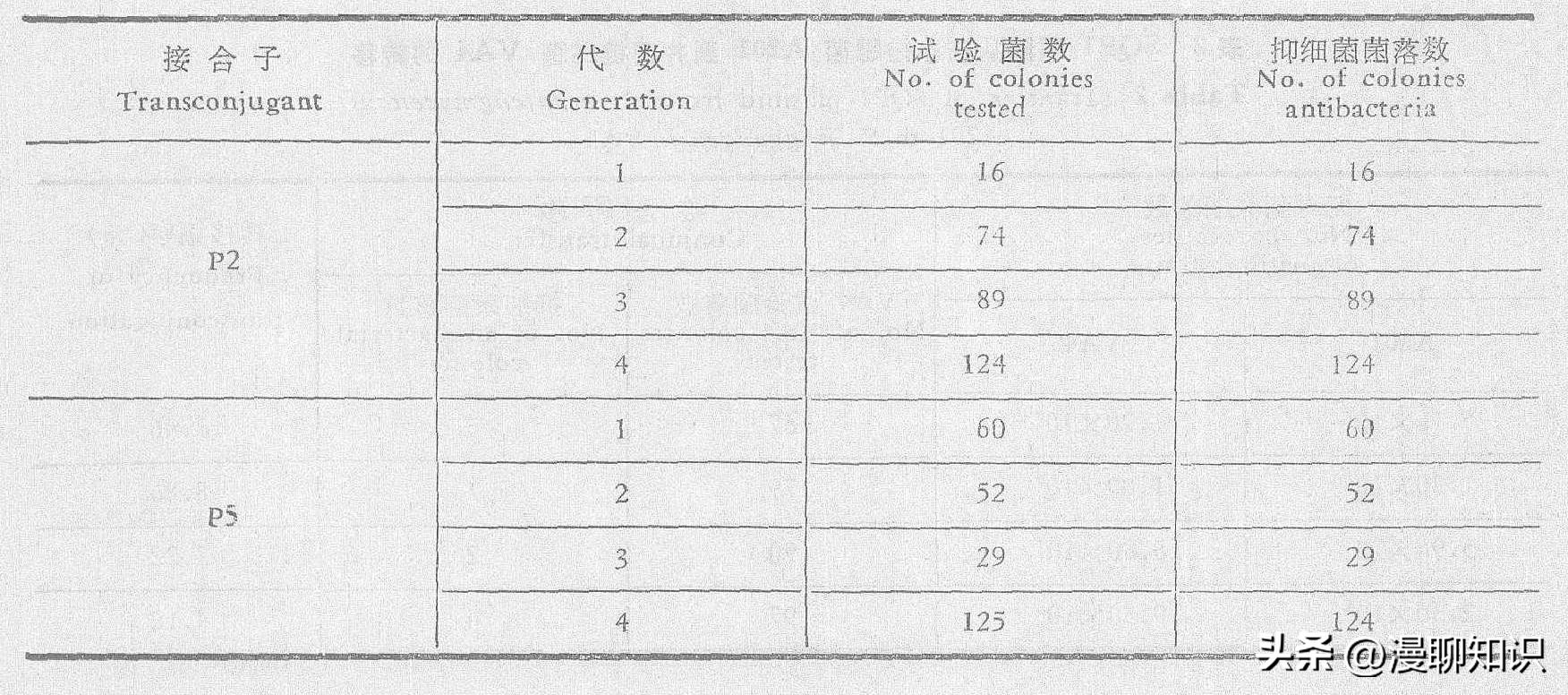
为了了解接合子抑制细菌活性的遗传稳定性,我们对接合子P2、P5菌株进行了连续单菌分离,每一代挑取一定数量的单菌落进行抑制细菌活性测定,结果指出,连续四代几乎没有分离现象,表明其遗传上是稳定的。
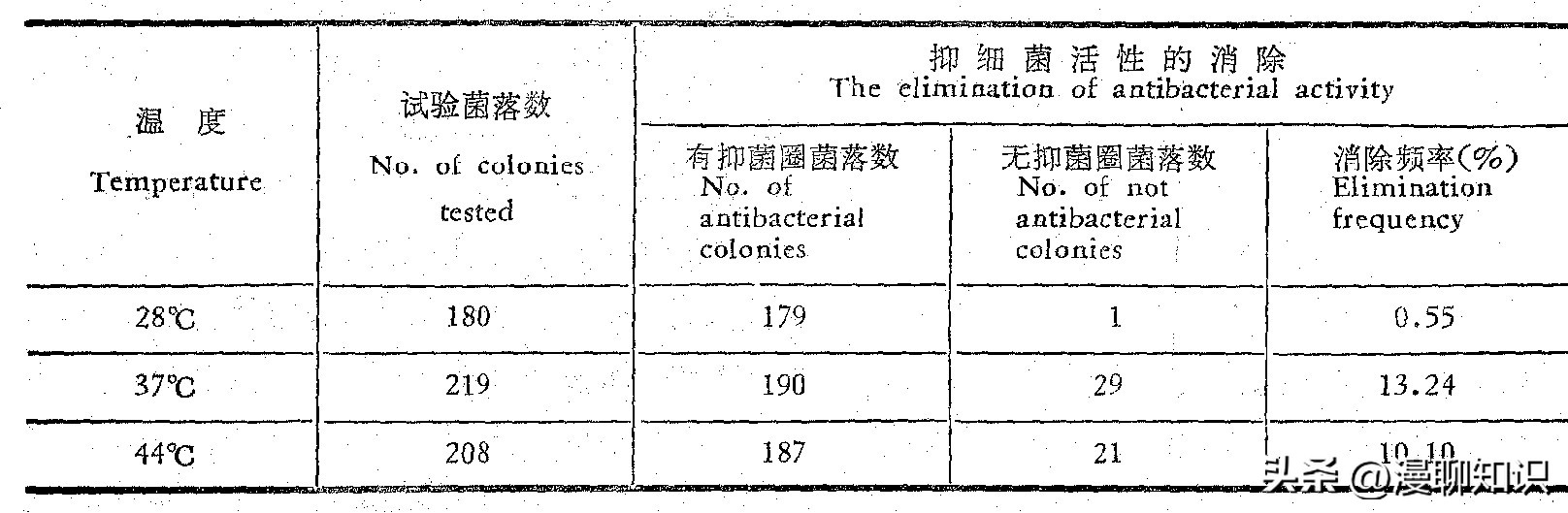
(四)高温培养对P2接合子抑制细菌活性的消除作用
如果接合子P2的抑制细菌活性是由于SQP1质粒接合转移的结果,那末已知的质粒消除因子将促进这种表型的消除,我们试验了高温预培养的影响,结果表明,37℃和44℃高温预培养使抑制细菌活性的消除频率都达10%以上,而28℃培养时的消除频率为0.55%,这和庆丰链霉菌原养型M15菌株SQP1质粒的自发消除频率相当。
表4 高温预培养对P2菌株抑细菌活性的消除
(五)接合子抑制细菌产物的鉴定
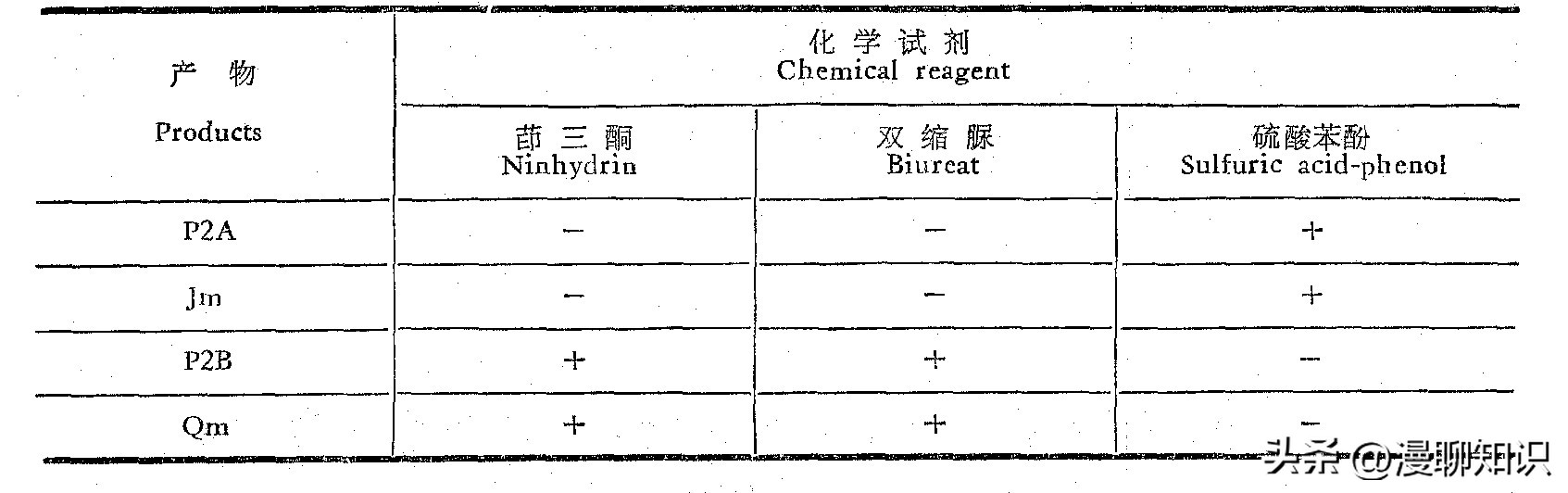
为了鉴定接合子P2所产生抑制细菌物质的性质,我们将P2菌株进行液体培养,并从培养液中按方法和材料中所述的步骤进行分离,结果表明,P2菌株能发酵产生二种物质,从分部收集的第2管到第30管为对硫酸苯酚反应为阳性的物质称为P2A,从40管以后则出现一个274毫微米波长处有紫外吸收的物质称为P2B,我们取P2A及P2B进行一系列性质测定。
紫外吸收谱: P2A具有末端吸收的特点,而P2B则为一典型的胞嘧啶核苷类化合物吸收谱,它在波长274毫微米处有一吸收峰,和庆丰霉素的紫外吸收谱一致。
薄层层析:经硫酸焦化显影,P2A斑点的Rf值为0.9,和Jm位置相同,P2B斑点的Rf值为0.26,和Qm位置相同,并在紫外线下显现萤光。

生物显影:P2A和Jm都无抑细菌活性,而P2B则和Qm一样能抑制枯草杆菌的生长。
抗菌谱测定:P2A及Jm不抑制所有试验菌,P2B.及Qm除了抑制枯草杆菌外还抑制八叠球菌、红酵母、大肠杆菌。
此外,P2A和Jm对茚三酮及双缩脲试剂呈阴性反应,而P2B及Qm除对硫酸苯酚试剂为阴性外对茚三酮及双缩脲试剂则呈阳性反应。
由此证明接合子产生的二个抗菌物质P2A为Jm,P2B为Qm。
按方法和材料中所述程序取得试验菌株发酵液,通过阳离子树脂Dowe×50 (Na+)交换柱,用蒸馏水洗柱后,以2N HCl洗脱,分部收集,每管为3毫升,逐管测定硫酸苯酚试剂显色反应及274毫微米波长处的光密度,合并硫酸苯酚阳性反应部份(P2A)及274毫微米吸收峰部分(P2B),分别通过阴离子树脂711 (OH-),使之中和,然后减压浓缩,于4℃贮存备用。
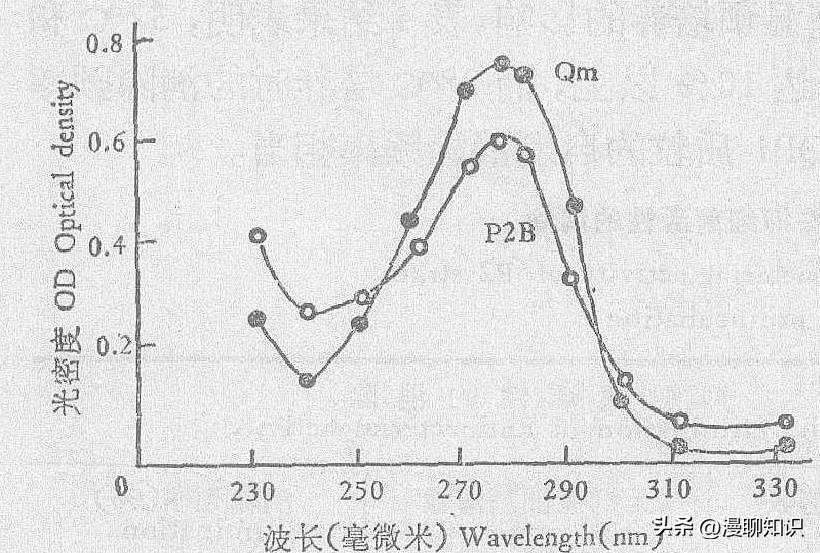
图5 P2B紫外吸收光谱
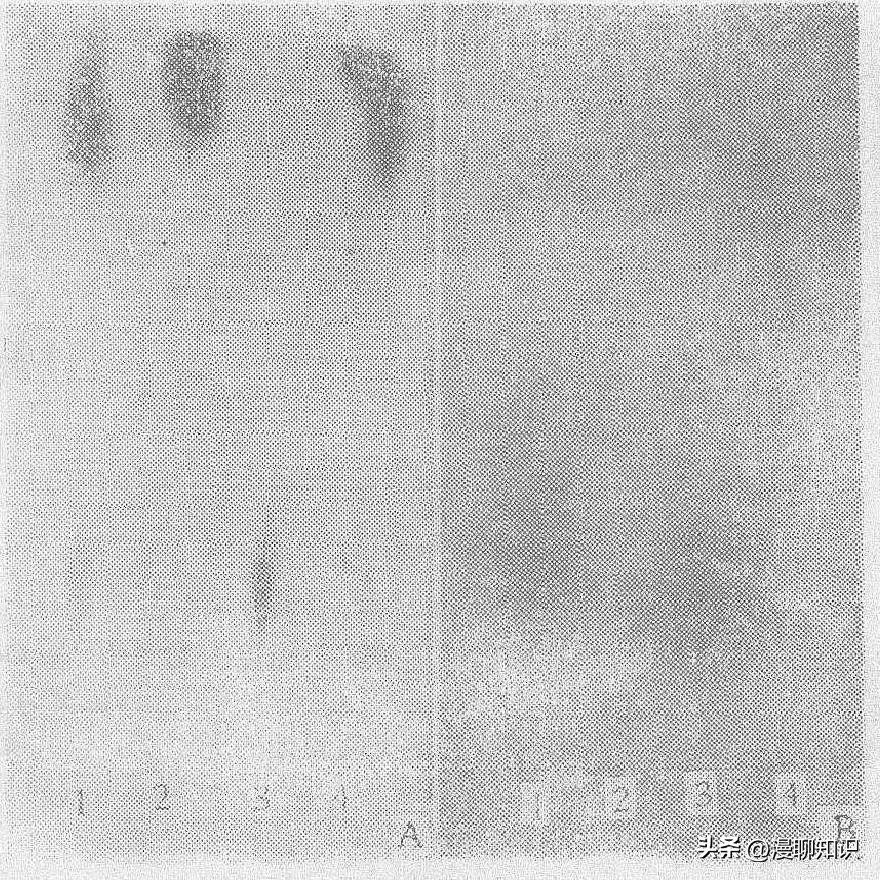
图6 P2A及P2B的薄层层析
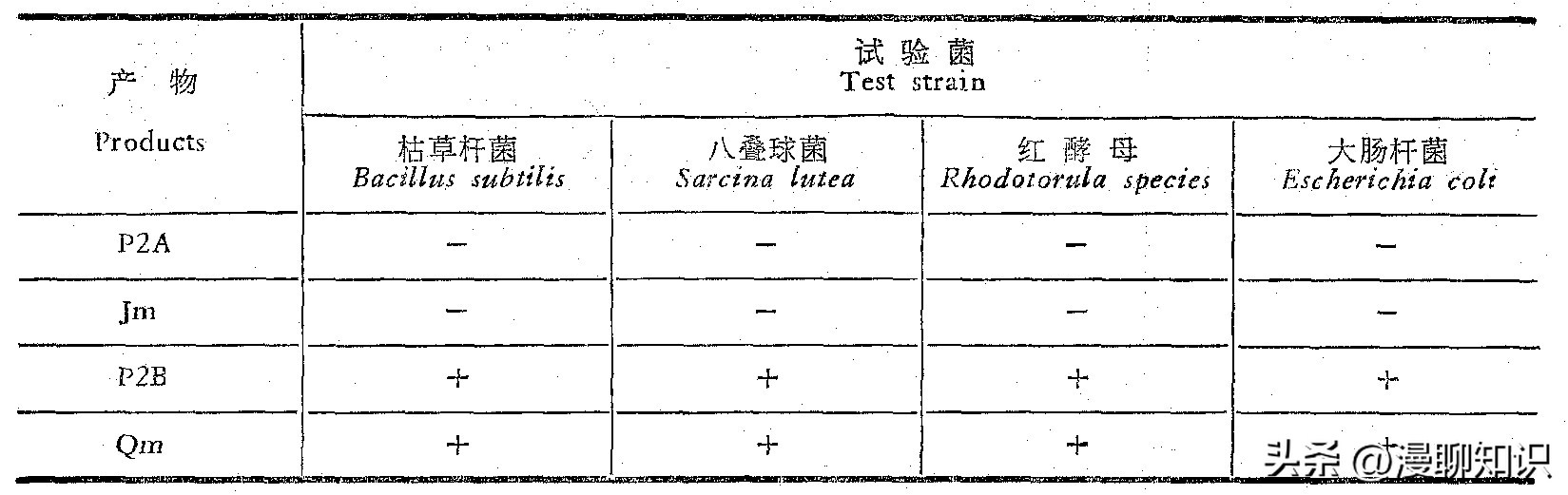
一:不抑制No inhibition,+:抑制Inhibition
讨论
以上的实验数据指出,庆丰链霉菌SQP1+的衍生菌株A201和相当于SQP1-的井冈链霉菌衍生菌株VA4经过混合培养后,SQP1质粒能够通过接合作用由前者转移给后者,使接合子获得了合成抑制细菌抗生素的能力,其转移频率为2—8%。
井冈链霉菌是井冈霉素(即有效霉素,validamycin)的产生菌,它不能抑制细菌,接合子所以能抑制细菌,是由于获得了SQP1质粒的缘故,我们对接合子发酵产物分离鉴定的结果表明,它产生的二个抗菌物质,P2A为Jm,P2B是Qm,而亲株VA4只产生Jm而不产生Qm,据此,我们推测SQP1质粒很可能含有庆丰霉素生物合成的基因,因之当SQP1质粒进入受体菌细胞,就能使它利用本身的初级代谢产物为原料而合成庆丰霉素,成为能同时产生Jm和Qm的菌株。
这是Hopwood报道了天蓝色链霉菌A3(2)的SCP1质粒具有种间转移,并编码次甲基霉素生物合成途径酶的第一个例子以来的又一个例子。
在自然界中存在着很多这样的例子,即属于完全不同的微生物种和属的菌株,往往可以产生化学结构完全相同或相似的抗生素,一个菌株又往往可以产生化学结构上完全不相关的几种抗生素,这可能也是质粒控制抗生素合成途径酶的结构基因,并通过接合作用而发生种间或属间转移的例证。

一个外源质粒要进入一个新的寄主细胞而稳定存在,会受到一些因素的限制,如质粒的不相容性,限制修饰系统等。
到现在为止,我们并不知道井冈霉素的生物合成是否有质粒参与,也不知道井冈链霉菌细胞内是否有别的质粒存在,如果有的话,那末它和SQP1质粒将属于不同的不相容性组。
至于链霉菌中的限制修饰系统,虽已有过一些报道[12,13],但在我们的实验中根据SQP1质粒能以比较高的比例进入细胞,并稳定表达这些结果,可能说明井冈链霉菌对SQP1质粒没有明显的限制作用。
《遗传学报》
《遗传》
《微生物学报》