
文/泊客
编/泊客
前言
沙眼衣原体是一种细菌性病原体,可引起 人类眼部和生殖器官感染 。
沙眼衣原体只在宿主细胞内的一个特征液泡内繁殖,在液泡内它通过向宿主细胞注射III型分泌效应蛋白来操纵宿主细胞。

在这里,我们确定 CteG 是第一个与高尔基体相关的沙眼衣原体效应体。
为此,我们构建了沙眼衣原体表达候选效应物融合到双血凝素(2HA)标签的菌株。
免疫荧光显微镜观察发现,CteG-2HA已进入感染细胞的细胞质。
感染后16-20 h, CteG-2HA多与高尔基体相关。

然而,CteG-2HA也出现在宿主细胞膜上,在感染后30或40 h,这是其主要的定位。
这种CteG-2HA主要定位的改变与完整的微丝或微管无关。
在未感染的细胞中异位表达CteG(656个氨基酸残基)的不同区域,发现其前100个残基包含一个高尔基靶向区。
虽然沙眼衣原体cteG突变体在细胞内增殖过程中没有表现出缺陷,但cteG在酿酒酵母中表达时却引起了蛋白的空泡分选缺陷。

这表明CteG可能通过破坏宿主细胞的囊泡转运发挥作用。
第一部分
一、CT105-2HA由沙眼衣原体输送到被感染细胞的细胞质中
为了检测候选衣原体T3S底物CT053、CT082、CT105、CT429和CT849是否可以被衣原体转移到宿主细胞的细胞质中,
沙眼衣原体c端编码有双血凝素(2HA)表位标签的这些蛋白的质粒被转化为沙眼衣原体L2/434。

通过免疫印迹法确认了含有编码CT053-2HA(预计分子质量为17 kDa)、CT082-2HA (60 kDa)、
CT105-2HA (68 kDa)、CT4292HA (39 kDa)或CT849-2HA (18 kDa)质粒的沙眼衣原体菌株感染HeLa细胞40 h后的蛋白生产。
产生CT053-2HA、CT082-2HA和CT105-2HA的菌株在SDS-PAGE上迁移的分子质量与预测的全长蛋白的分子质量不同,
正如之前观察到的,在Y. enterocolitica中产生了相同的2ha标记蛋白版本。

总之,这些实验证实了构建的 沙眼衣原体菌株表达了预期的2ha表位标记蛋白 。
为了分析CT053-2HA、CT082-2HA、CT105-2HA、CT429-2HA和CT849-2HA的亚细胞定位,
我们用携带相应编码质粒的沙眼衣原体l2 /434源株感染HeLa细胞20或40 h,然后用免疫荧光显微镜分析。

在20 p.i.或40 h p.i., CT053-2HA、CT082-2HA、CT429-2HA和CT849-2HA仅在与沙眼衣原体主要外膜蛋白(MOMP)信号共定位中被检测到。
相反,CT105-2HA在包涵体外被检测到,表明它是通过细菌传递到受感染的宿主细胞的细胞质中。
二、CT105的表达主要局限于沙眼衣原体LGV菌株
CT105的全长同源基因仅存在于C. muridarum和C. suis中,CT105的表达主要局限于沙眼衣原体LGV菌株。

CT105 是一种含有656个氨基酸残基的蛋白,其序列与其他衣原体中可能存在同源序列的蛋白没有显著的相似性,
但CT105全长同源序列仅在C. muridarum和C. suis中发现。
在猪链球菌和其他衣原体中,不同的开放阅读框可能编码的蛋白质仅与CT105的部分氨基酸序列具有一定的同源性(30-22%)。
如前所述,通过 反转录定量PCR (RT-qPCR) 对不同沙眼衣原体菌株中ct105 mRNA水平的分析表明,该基因仅在LGV菌株(血清L1-L3)中显著表达。

此外,正如前面提到的,考虑到ct105启动子区基于沙眼衣原体LGV菌株L2b/UCH-1/proctitis中确定的转录起始位点,
沙眼衣原体非LGV菌株(serovars A-K)在沙眼衣原体(沙眼衣原体)δ 66识别的−10区域上游缺少74个核苷酸。
此前也有研究表明,ct105是眼沙眼衣原体(c - c)株的假基因。
因此,在沙眼衣原体血清中,活性CT105主要由LGV菌株产生。

三、表达质粒编码CT105-2HA的沙眼梭菌的特性
将含有pCT105-2HA的L2/434菌株与亲本菌株进行比较。
两株沙眼衣原体发育周期中ct105的 表达谱相似。
与先前的观察结果一致, ct105 mRNA的最高水平在2小时p.i被检测到。

此外,在2 h p.i.携带pCT105-2HA的菌株显示ct105 mRNA水平比亲本菌株增加了10倍。
然而,质粒编码CT105-2HA在沙眼衣原体中的表达对衣原体生长没有显著影响。
在携带pCT105-2HA的沙眼衣原体L2/434感染的HeLa细胞的提取物中,免疫印迹可在16 ~ 40 h p.i检测到2ha标记的蛋白。

除了可能与全长CT105-2HA (68 kDa)相对应的条带外,在20至40 h p.i之间一致观察到一些分子质量较低的更快迁移物种。
通过免疫印迹法,我们也观察到了类似的结果。
免疫印迹法 检测了HeLa细胞被携带质粒(Ptet -CT105-2HA)的L2/434感染后的提取液,在质粒中,CT105-2HA的产生是由四环素启动子(Ptet)驱动的。

这表明质粒编码的CT105-2HA随着沙眼衣原体发育周期的进展而被修饰/降解。
对含有pCT105-2HA或pTet-CT105-2HA的L2/434感染的HeLa细胞的提取物,
或相应的富含衣原体的提取物的免疫印迹表明,CT105-2HA的修饰/降解发生在细菌内部。

四、CT105-2HA定位于沙眼梭菌感染细胞的高尔基复合体
为了进一步分析CT105-2HA在 HeLa细胞感染过程中的定位 ,
我们用含pCT105-2HA的沙眼衣原体L2/434对HeLa细胞进行不同时间的感染,然后用免疫荧光显微镜进行分析。
在2,4,8 h p.i.中,CT105-2HA可检测到与沙眼衣原体Hsp60信号共定位,表明CT105-2HA存在于细菌细胞内。

在16和20小时时,CT105-2HA分别在约37%和90%的感染细胞的包涵体外被检测到;
在30和40 h / i时,在所有感染细胞的包涵体之外都发现了该蛋白。
而在16、20和30 h p.i.时, CT105-2HA在宿主细胞的细胞质中被检测到 ,
CT105-2HA蛋白集中在包涵体的一侧,在30 h p.i.时,该蛋白也在被感染细胞的外围被检测到。
此外,在40h p.i时,CT105-2HA主要出现在感染细胞的周围。

CT105-2HA在被感染细胞胞浆内的包涵体附近积累,提示其可能 定位于高尔基复合体 。
CT105-2HA的免疫荧光信号在较长时间感染时也不那么紧密,这使人联想到沙眼衣原体感染细胞中的高尔基体碎片。
为了分析这一点,将携带pCT105-2HA的沙眼衣原体L2/434感染的HeLa细胞固定在16、20、30或40 h p.i.,然后用免疫荧光显微镜进行分析。
这表明,CT105-2HA在16、20和30小时p.i时定位于高尔基区,而在40小时p.i时则不那么明显。

CT105-2HA与顺式高尔基(GM130)和TGN (TGN46)标记物没有完美的共定位。
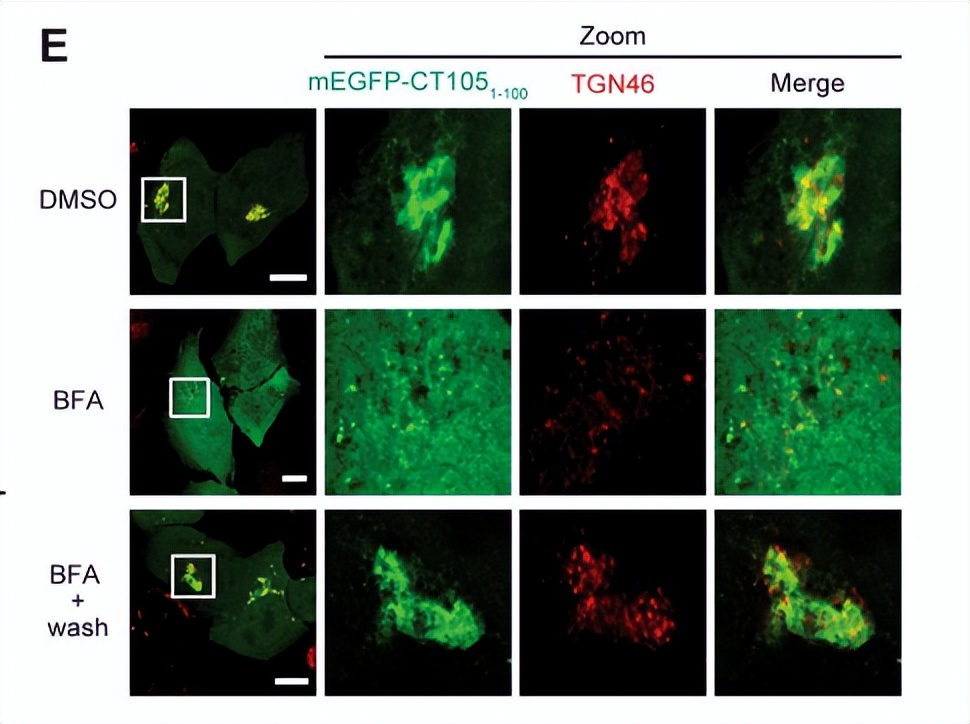
为了进一步证实CT105-2HA与高尔基体的相关性,用含pCT105-2HA的L2/434株感染HeLa细胞后,用Brefeldin A (BFA)处理细胞,导致 可逆的高尔基体碎裂 。
为此,在19小时p.i.,感染细胞或与BFA或单独二甲基*砜亚**(DMSO)溶剂孵育1小时,然后固定。

将感染细胞与BFA孵育1 h,从BFA中冲洗出来,孵育1 h后固定,免疫荧光显微镜显示,
在dmso处理的感染细胞中,CT105-2HA在高尔基区积累,与未经处理的细胞感染16或20小时p.i相似。
gm130标记的高尔基体破碎成小泡,CT105-2HA的免疫荧光信号分散,且大多低于检测限。
在经BFA处理后被药物洗出的细胞中,gm130标记的高尔基体和CT105-2HA的免疫荧光信号大多致密且靠近包膜。

这表明,在最初检测到CT105-2HA由沙眼衣原体输送到受感染的宿主细胞的细胞质时,CT105-2HA在16-20小时p.i.时与高尔基体结合。
第二部分
我们发现沙眼衣原体CT105蛋白被传递到被感染的宿主细胞的细胞质中。
在衣原体发育周期的不同时期,CT105在高尔基复合体(16-30 h / i)/在宿主细胞膜(30-40 h / i.)聚集。

由于CT105可由Y. enterocolitica的T3S系统分泌,并可干扰啤酒酵母的囊泡运输,
这表明CT105是一种新发现的沙眼衣原体T3S效应因子。
然而,到目前为止,我们还没有直接证据表明CT105能够干扰被感染的人类细胞的囊泡贩运,这必须在未来的研究中进行直接分析。
总的来说,这项工作 扩大了已知的衣原体效应蛋白的组合, 并揭示了第一个与高尔基复合体相关的衣原体蛋白。

此外,它说明了单个沙眼衣原体效应者在衣原体发育周期中,可能具有不同的亚细胞定位和不同的功能。
在感染过程中,这种双重定位可能是一种多样化效应器功能的手段,它唤起了沙门氏菌的SopB效应器,
该效应器最初定位在宿主细胞膜上以介导细菌入侵,但随后被定向到含沙门氏菌的液泡中,以调节细菌的细胞内生长。

除了CT105,沙眼衣原体效应物CT867,一种被称为ChlaDub216或Cdu217的去泛素酶,在不同的感染时间在包膜和宿主细胞膜上被检测到。
CT105和CT867/ChlaDub2/Cdu2在感染后期的定位可能表明其在宿主细胞出口中的作用。
CT105和CT867/ChlaDub2/Cdu2如何区别地定向到不同的宿主细胞定位尚不清楚。
在CT105的情况下, 不太可能涉及囊泡贩运, 因为微丝和微管的破坏并不影响其在感染细胞的质膜上的定位。

我们设想,宿主细胞质CT105通过共价修饰/宿主细胞膜脂质/蛋白质组成的改变,特异性地指向不同的亚细胞定位。
在感染过程中,细菌效应蛋白精确的空间和时间亚细胞靶向是其功能的基本特征。
在其他细菌病原体中,至少有三种其他效应蛋白已被证明定位于感染宿主细胞的高尔基体。
这些是大肠杆菌的EspI/NleA,嗜肺L. pneumophila49的GobX和肠沙门氏菌的SteD。
EspI/NleA干扰coatomer蛋白II复合物功能,抑制宿主细胞蛋白分泌,并破坏肠道紧密连接。
此外, EspI/NleA可抑制NLRP3炎症小体的激活 。
GobX显示E3泛素连接酶活性,但这如何促进嗜肺L.肺炎杆菌感染尚不清楚。
最后,SteD干扰宿主E3泛素连接酶的活性,导致表面定位的主要组织相容性复合体II类分子的减少和T细胞活化的抑制。

这说明CT105不能根据其在高尔基的定位来预测其特定的功能。
当CT105蛋白在Hela细胞中异位表达时,CT105蛋白的前100个氨基酸残基足以定位高尔基体。
然而,这一区域并没有在其他相关的真核或细菌蛋白中发现明显的基序,CT105高尔基体定位的确切分子决定因素仍有待确定。
其他相关的问题是,是否同样的决定因素在感染细胞中负责高尔基体定位,以及全长CT105蛋白的哪些特性使其能够靶向质膜。

其他定位于感染细胞高尔基体的效应蛋白使用不同的机制将它们导向细胞器。
在EspI/NleA中,高 尔基体的靶向部分 是由其c -末端的突触后密度95/椎间盘大/咬合小带-1 (PDZ)域介导的。
CT105也有一个假定的PDZ domain56,但在哺乳动物细胞中异位表达后,相关残基不参与其亚细胞定位(数据未显示)。
CT105在沙眼衣原体血清中表现出显著的变异性,揭示了lgv特异性的遗传和转录组特征。

这表明CT105可能具有 与这些菌株感染的独特特征相关的特定功能 ,
与眼部和泌尿生殖道沙眼衣原体菌株不同,CT105可以感染单核吞噬细胞并传播到区域淋巴结。
这也表明LGV株感染的特征应该是一种维持ct105基因活性的进化压力。
此外,CT105的全长同源体仅存在于C. muridarum和C. suis中,这与这两种衣原体是沙眼C.沙眼C. 3的*亲近**一致。

综上所述,我们确定 CT105是沙眼衣原体的T3S效应因子,
它定位于感染细胞的高尔基体和质膜,并可以破坏向酿酒酵母液泡的运输。
由于CT105是第一个与高尔基复合体相关的衣原体效应因子,我们建议将其命名为CteG(与高尔基复合体相关的沙眼衣原体效应因子)。

CteG的鉴定也留下了许多有待解决的问题,包括其功能、亚细胞靶向机制、多样性和在沙眼衣原体和衣原体中的特异性。
参考文献:
1.Borges, V. & Gomes, J,《利用小肠结肠炎耶尔森菌作为异种系统鉴定沙眼衣原体III型分泌底物》
2.Albrecht, M., Sharma, C. M,《对沙眼衣原体淋巴肉芽肿性病分离株的深度比较基因组学强调了可能涉及发病适应的基因》
3.Heuer, D.,《沙眼衣原体ⅲ型分泌伴侣-底物复合物的鉴定》
4.Shohdy, N., Efe, J. A.,《基于深度测序发现沙眼衣原体转录组》