
前言
百喜草是一种形成土壤的、根茎状的暖季多年生草,广泛生长于美国东南部和中南美洲。
百喜草耐低土壤肥力、干旱、短期洪水和密集放牧,非常适合低投入系统。
百喜草耐多种落叶制度,可用于各种放养方法。
在佛罗里达州,暖季多年生牧场和牧场占地约220万公顷,供养着168万头牛。

百喜草占据了该地区超过100万公顷的面积,为佛罗里达州大约100万头奶牛提供了放牧饲料。
通过植物引种和育种工作,已经开发出了几个百喜草品种,其中“Pensacola”是种植最广泛的。
彭萨科拉是一种相对卧二倍体类型,叶片窄,种子产量好,在集约放牧下具有持久性。
已经开发的其他百喜草品种表现出截然不同的生长特性。

“UF Riata”是从涉及彭萨科拉的杂交中选择的一个品种,其目标是鉴定一个对日长不太敏感的品种,以增加早春和早秋的饲料产量并延长放牧季节。
UF Riata的生长习性也比Pensacola更为直立,类似于“Tifton 9”百喜草。
放牧强度是决定饲料产量、营养价值和植物持久性的关键。将放牧间隔从2周增加到4周,包括UF Riata在内的几个百喜草品种的年牧草积累(HA)增加了约50%。
此外,与隔4周放牧相比,隔2周放牧的UF Riata的盖度和根茎质量更小,这表明该品种更适应较少的落叶。

Gates et al.(1999)指出,当百海草在2年内每两周放牧一次时,黄化春季生长明显减少,这表明可供生长的资源枯竭。
Vendramini等人(2013)认为,降低直立生长的百喜草品种的放牧频率是提高HA和持久性的有效管理措施。
关于UF Riata百喜草的一个问题是,较大的冷季产量是否会影响其长期持久性和生产力,以及生长季节不同程度的落叶频率和强度如何影响其长期持久性。
为了回答这个问题,Interrante等人(2009)评估了两种茬高(4和8厘米)和两种收获频率(7和21天)对五种百喜草的影响。

作者指出,每21天采收一次,残茬高度达到8cm时,红柳的覆盖面积最大,而频繁的落叶导致红柳的覆盖面积最小。
相比之下,“阿根廷”百喜草的覆盖不受落叶制度的影响。
此外,UF Riata的根茎质量、茎基氮(N)和总非结构性碳水化合物含量低于阿根廷和Pensacola。
因此,百喜草的落叶管理可能比其他品种更为关键。

然而,放牧和修剪研究可能会产生不同的结果和推论,虽然之前已经对UF Riata百喜草的放牧频率进行了研究,但没有关于放牧强度如何影响其对草地反应的信息。
通过同时提高载畜率(SR)和氮肥水平来提高牧场管理的总体强度,对彭sacola bahi禾草牧场HA率、粗蛋白质(CP)和体外可消化有机质(IVDOM)浓度有积极影响。
例如,随着管理强度从低到高, HA率、CP和IVDOM浓度分别从17至41公斤公顷−1天−1、99至140克kg−1和459至505克kg−1。
在连续放养条件下,在相同施氮量和不同施氮量的情况下,随着施氮量的增加,百叶草牧草质量(HM)和牧草补给量呈线性下降,而CP和IVDOM浓度随着施氮量从4 ~ 12头牛/ ha−1的增加呈线性增加。

但SR与HA率呈二次关系。研究人员发现,在SR为8头小牛HA - 1时,HA率最高,残茬高度约为30厘米。
这个高度提供了足够的叶面积来优化光拦截,从而提高了HA率。
相比之下,12头母牛/公顷的牧场残茬高度约为10厘米,这减少了叶面积,最终降低了ha率。
相反,每公顷放养4头小牛的牧场会产生过多的HM,导致自遮荫、衰老加剧,随后ha率降低。

因此,我们可以推断,当施氮量增加时,百海草的HA率会随着SR的增加而增加,但当不提供这样的N供应时,增加SR超过一定水平可能会降低HA率。
Hirata等人(2006)综合了五次百喜草放牧试验的数据,并报告称,通过干物质(DM)消化率和粗蛋白质浓度测量,氮肥施用量的增加导致HA率的提高和营养价值的改善。
然而,他们也观察到,随着HM的增加,DM消化率和粗蛋白质浓度降低。
本研究研究了相同施氮量下不同放牧强度对百喜草的影响。

虽然SR是放牧管理中的一个重要因素,但它对动物生产性能的影响是间接的,因为它不考虑冠层的质量和数量。
因此,不同的放牧强度是基于施加不同程度的放牧后残余HM。
到目前为止,还没有研究使用不同水平的放牧后残留HM来评估UF Riata百喜草的换草反应。
本研究的目的是评价百喜草对不同放牧强度下的草地响应;包括HA、营养价值、根茎质量、土壤覆盖度和杂草比例。

基于放牧后剩余HM施加了3种不同的放牧强度。
我们推测,增加放牧强度会提高百喜草的饲料营养价值,但会降低总HA和HA率、根际生物量和土壤盖度,从而增加杂草对草地的侵蚀。

材料与方法
实验场地
该实验于2018年、2019年和2020年在佛罗里达州马里安娜北佛罗里达研究与教育中心、佛罗里达大学食品与农业科学研究所进行。
实验场地的土壤是红湾细砂壤土。在试验开始之前,研究地点是一个建立了大约10年的根茎花生田。
2017年,使用麦草畏(N,N-二-(3-氨基丙基)甲胺盐3,6-二氯-o-茴香酸)和农达PowerMAX II(草甘膦,N-(磷甲乙基)甘氨酸,以其钾盐的形式)杀死了根状花生。
2017年4月11日,麦草畏和草甘膦的施用量分别为0.38和0.77 L ha−1。此外,在2017年4月25日和2017年5月11日,草甘膦的施用量为1.91 L ha−1。

试验期间的天气资料见表1。土壤样本收集在2017年5月报道pHwater 6.1, 7.9 g公斤−1土壤有机质(OM)和Mehlich-1-extractable P = 12.5毫克公斤−1(低),K = 36.5毫克公斤−1(媒介),mg = 57.5毫克公斤−1(媒介),Ca = 358毫克公斤−1(足够的),S = 10毫克公斤−1(低),B = 0.16毫克公斤−1(低),锌= 0.9毫克公斤−1(低),Mn = 30.5毫克公斤−1(非常高),铁= 8毫克公斤−1(足够的),和铜= 0.25毫克公斤−1(低)。
本研究是在包括棉花在内的其他项目的相同一般实验区进行的。由于这个原因,灌溉管理遵循佛罗里达大学/食品和农业科学研究所对棉花的标准推广建议。2019年和2020年灌溉水数据如表1所示。

虽然没有提供2018年的数据,但由于2018年降雨分布较好,灌溉用水的使用频率较低。分别于2017年12月1日、2018年11月20日和2019年11月25日在试验田播种56 kg ha - 1的“佛罗里达401”黑麦和“RAM”燕麦,作为冬季牧草。
在一年中的这个时候过度播种百喜草是可能的,因为它的休眠是由白天的长度和冬天的温度下降引发的。
冷季牧草没有使用任何机械或化学方法终止,放牧从冷季过渡到暖季,因为冷季牧草结束了它们的循环(自然终止),百喜草开始活跃生长。
关于地块冷季管理的更多信息可以在Santos等人(2023)中找到。
表1。北佛罗里达研究与教育中心(NFREC) Marianna, FL,在实验年份的每月天气状况。

治疗、实验设计和动物护理
2017年5月11日,UF Riata百喜草在7.3米× 15.2米的地块上以28公斤种子公顷的面积进行了钻探。
2018年开始放牧时,这些地块已经完全建立起来。施用3种放牧强度,分别为重度放牧(HG)、中度放牧(MG)和轻度放牧(LG)。
放牧强度以牛为对象,目标放牧后HG、MG和LG的剩余HM分别为500、1500和2500 kg DM ha - 1。

当HM低于1000-1500 kg DM / 1时,放牧动物的牧草采食量会大幅下降。
因此,放牧后HM选择1500 kg DM ha - 1来模拟MG系统,放牧后HM分别选择500和2500 kg DM ha - 1来模拟HG和LG系统。
处理采用完全随机区组设计,设3个重复。佛罗里达大学机构动物护理和使用委员会批准了本研究中使用动物的所有程序。

实验管理
试验田用电围栏隔开,并有单独的水槽。每年施两次肥,第一次施氮17公斤,磷肥22公斤,钾肥83公斤,第二次施氮72公斤。
第一次和第二次施肥分别发生在2018年6月8日和6月25日,2019年6月11日和7月31日,2020年5月28日和6月19日。
按照土壤试验分析建议施肥。每年应用0.15 L ha - 1(商业配方)的Engenia 于2018年5月31日、2019年5月29日和2020年6月12日进行检测。

大约每14天用安格斯小母牛和泽西阉牛对地块进行暴民放养。
2周的频率间隔旨在模拟目标茬高保持不变的放牧系统。
2018年、2019年和2020年放牧季初实验动物平均体重(kg)和标准差分别为508±30、445±41和518±30。
放牧前HM大于目标放牧后HM时发生放牧。
为避免环境温度过高,放牧时间分别为0430 - 1100 h和1630 - 2000 h。

在白天较热的时间,动物被带到实验地块旁边的休息畜栏,在那里它们可以获得阴凉处和水。
根据环境温度和放牧前HM的不同,整个试验区的放牧时间在1天以上,但不超过2天。
同一小区内所有试验单位(小区)均在同一天放牧。
在放牧期间,动物们在一个保留的百喜草牧场上。
在放牧活动的前一天晚上,动物被带到休息的畜栏并禁食12小时。

2018年4月24日、2019年4月30日和2020年5月27日是百喜草每年的第一次放牧事件。
受新冠肺炎疫情限制,2020年放牧推迟,并于2020年5月5日以7.5 cm茬高的刈割方式分田。
2018年共有13个放牧周期,2019年12个,2020年11个。
考虑到冷季放牧向暖季放牧的过渡时期,每年仅纳入11个放牧周期进行统计分析。每个放牧周期的具体日期见表2。
表2。放牧周期和每年的放牧日期。

双采样技术(Haydock & Shaw, 1975)用于估计放牧前后的HM。
该技术包括将0.25平方米样方收获的饲料质量回归到铝盘沉降高度(间接测量)上的土壤水平(直接测量)。
每28天使用24个配对样本进行新的校准。
放牧前,采用圆盘沉降高度(每地块30点)间接测量放牧前HM。

放牧期间还测量了盘沉降高度。一旦达到目标高度,将动物移出地块并记录放牧后的HM。
两个动物在每个地块开始放牧,当它们在原地块达到放牧后HM目标时,它们可以重新分配到新的地块。
不同年份和放牧周期,HG、MG和LG的平均沉降高度分别为2.4 (0 ~ 4.6 cm,范围)、6.5(3.9 ~ 12.2,范围)和10.8 cm (7.3 ~ 20.4 cm,范围)。
方程的平均R2为0.79,范围为0.45 ~ 0.96,在第一个放牧周期出现最小值。

响应变量
牧草响应
牧草的响应包括放牧前后HM、总HA、HA率、IVDOM和CP浓度、土壤覆盖度、非种植草百分比、其他杂草百分比和总杂草百分比。
放牧前和放牧后的HM是使用上述双重抽样技术确定的。
利用当前放牧周期放牧前HM与前一个放牧周期放牧后HM的差值估算总HA积累量,然后将所有放牧周期的值相加。
由于第一个放牧周期没有先前的放牧后HM,因此其价值被添加到总数中。
每年5个放牧周期的HA率由当前放牧周期的放牧前HM与前一个放牧周期的放牧后HM之差除以放牧周期之间的天数来确定。每个HA率的测量日期见表3。
表3。用于估算牧草积累率的周期和相应日期。

在每次放牧之前收集代表消耗的草料的手采草料样品,并用于确定营养价值。
样品在55°C的强制空气烘箱中干燥至恒重。
干燥的样品被磨碎,通过2毫米的筛网,使用威利磨机。
IVDOM是根据Moore和Mott(1974)描述的程序确定的。
为了测定N,使用混合机中的球磨机在25 Hz下运行9分钟,将2mm磨碎的子样品还原为细粉。
然后使用元素分析仪通过干燃烧分析样品的总N。
N浓度乘以6.25得到CP浓度。

杂草比例和土壤覆盖率
在每年最后一个放牧周期结束后,测定杂草比例和土壤覆盖度。
采用t Mannetje和Haydock(1963)描述的干重等级法确定杂草比例。
简单地说,在每个样地的20个随机点放置0.25 m2的金属环,现有植物种类排名第一、第二和第三。
如果物种在采样点的比例相似,则给予两者相同的等级。

这些比例乘以t Mannetje和Haydock(1963)提出的系数,分别为70.2、21.1和8.7,并相加得到各树种的干重比例。
杂草以三种不同的方式报告:未种植的杂草%,其他杂草%和总杂草%,即其他两者的总和。
非种植的禾草主要包括禾草,野生杂草和常见的百慕大草。
非草杂草以茶草和藜草为主。为了估计土壤覆盖总量,在量化杂草比例的相同20个点上进行了目测评估。
估算每个点的土壤覆被比例,然后取平均值,得到每个试验单元的单一值。

Root-rhizome生物量
根据Santos等人(2019)描述的方法,在每年最后一个放牧周期后测量根茎生物量。
简单地说,使用高尔夫球洞切割器从每个地块收集3个0至20厘米深× 10.8厘米直径的土芯。
将土芯(根+根茎+土壤)在55℃的强制空气干燥箱中干燥至恒重,并记录其重量。
土壤芯在850 μm筛子中洗涤,以去除土壤和异物,并反复从容器中倒入筛子。
除土后,根和根茎用相同的方法干燥。
记录单位土壤质量的根茎质量。每公顷根茎质量是根据2020年10月10日使用未扰动核心法确定的0至20厘米深度土层和体积密度估算的。
使用威利磨机研磨样品通过2毫米筛网。地面亚样品在600℃的马弗炉中燃烧5 h,以OM为基础表达根茎质量。

统计分析
数据分析使用SAS 9.4版的PROC GLIMMIX。
使用学生面板和PROC单变量检查所有变量的残差是否为正态性和方差齐性。
利用最小修正的赤池信息准则选择各变量的最佳协方差矩阵。
放牧前后HM和牧草IVDOM和CP浓度,放牧强度、放牧周期和年份为固定效应,阻滞效应为随机效应。

对于HA率,放牧强度、周期和年份被认为是固定效应,而块被认为是随机效应。
放牧周期和放牧期作为重复测量进行分析,根据PDIFF选项,LSMEANS差异有统计学意义(p < 0.05)。
土壤覆盖度、总杂草%、未种植草%、其他杂草%和根茎生物量以处理和年份为固定效应,以块为随机效应,以年份为重复测量。
由于这类数据的高度可变性,根据PDIFF选项,LSMEANS在p < 0.10时被认为具有统计学差异。
利用线性对比和二次对比评价了不同处理对土壤覆盖度、总杂草百分比、未种植草百分比、其他杂草百分比和根茎生物量的影响。根据f检验,p < 0.10为显著性差异。

结果
牧草响应
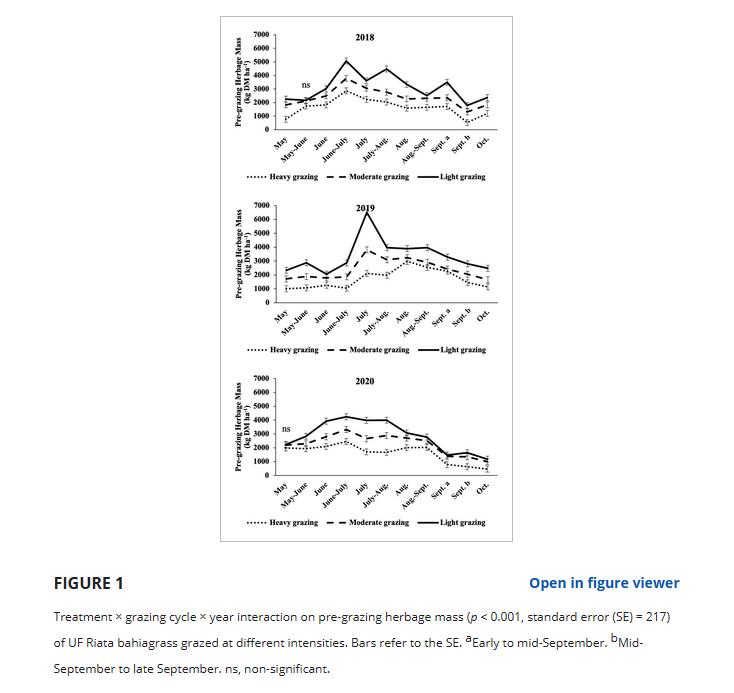
放牧前HM存在放牧强度×放牧周期×年的交互作用(p < 0.001, SE = 217)。
除2020年5 - 6月和2021年5月出现少数例外(处理间无差异)外,LG放牧前HM始终高于HG, MG处于中间状态(图1)。
2018年6 - 9月、2019年6 - 7 - 10月、2020年6 - 8月差异更为突出。
2018年和2020年放牧前土壤肥力最高的季节为6 - 7月,分别为5070和4040 kg DM ha - 1。
2019年,放牧前HM最大值出现在7月,LG为6020 kg DM ha - 1。
2018年和2020年放牧前土壤肥力最小,分别发生在生长季末的9月和10月,平均500 kg DM ha - 1。
2019年,放牧前HM最少发生在生长季开始的5月(1010 kg DM ha−1)。
放牧后HM存在放牧强度×放牧周期×年的交互作用(图2;p < 0.001, SE = 100)。
2018年和2019年,不同放牧周期的差异是一致的;放牧后HM以LG最高,MG次之,HG次之。
这种差异在2020年也基本一致,几乎没有例外。
2020年5 - 6月,MG和HG之间没有差异。
相反,2020年9 - 10月MG和LG之间没有差异。
在放牧周期和放牧年限中,放牧后LG、MG和HG的HM分别为2200、1480和780 kg DM ha - 1 (p < 0.001)。
尽管与目标HM(分别为LG、MG和HG,分别为2500、1500和500 kg DM ha - 1)有所不同,但在整个放牧周期和年份中观察到的放牧后HM水平确保了放牧强度的差异。

总HA没有处理(p= 0.49)或年份(p= 0.19)的影响,处理之间的平均值为11030 kg DM HA−1年−1。
不同放牧强度间HA率差异不显著,但受放牧周期×年交互作用的影响(p < 0.001, SE = 9.3;图3).不同年份之间的差异发生在整个放牧周期。
在2018年和2020年,6 - 7月的HA率最高,但2020年的HA比2018年高出约60%(分别为132和82 kg DM HA - 1 day - 1)。
2019年,HA最大发生在8 - 9月(112 kg DM HA−1 day−1),但7 - 8月差异不显著(108 kg DM HA−1 day−1)。
多年来,最低的HA率发生在生长末期,即9 - 10月。平均而言,2018年和2019年9 - 10月HA率为43 kg DM HA−1 day−1,显著高于2020年同期的13 kg DM HA−1 day−1。

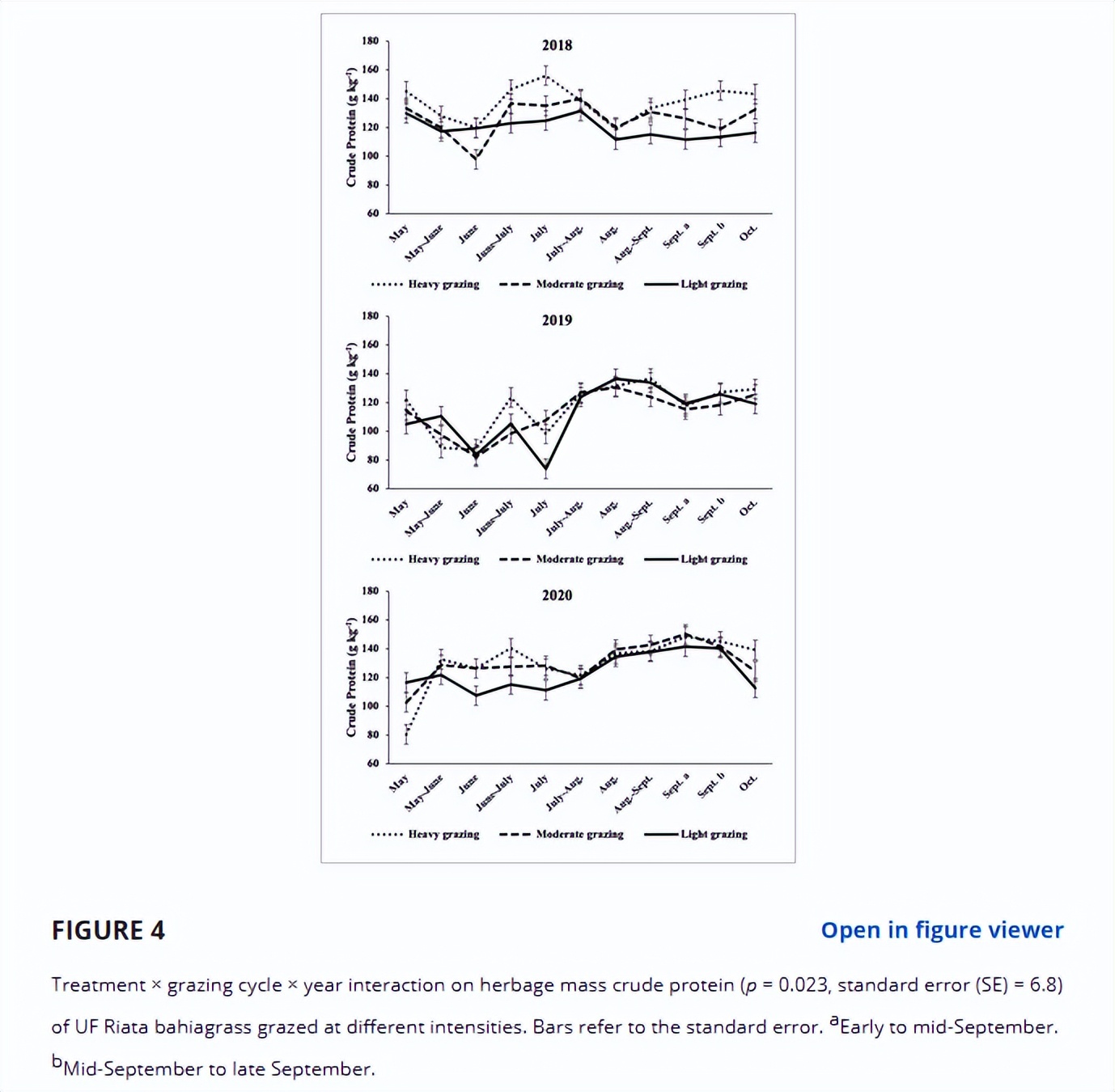
CP浓度存在处理×放牧周期×年的交互作用(p = 0.023, SE = 6.8)。
2018年、2019年和2020年11个放牧周期中,处理差异分别为55.4%、27.3%和36.4%。
除了少数情况,特别是在一年的早期放牧周期,HG导致CP浓度高于LG, MG处于中间位置(图4)。
2018年,差异发生在6 - 7月和9月(9月初)- 10月。
在2019年,这种差异发生在5月至6月、6月至7月和7月。
2020年5月、6月、6 - 7月和10月处理间存在差异。
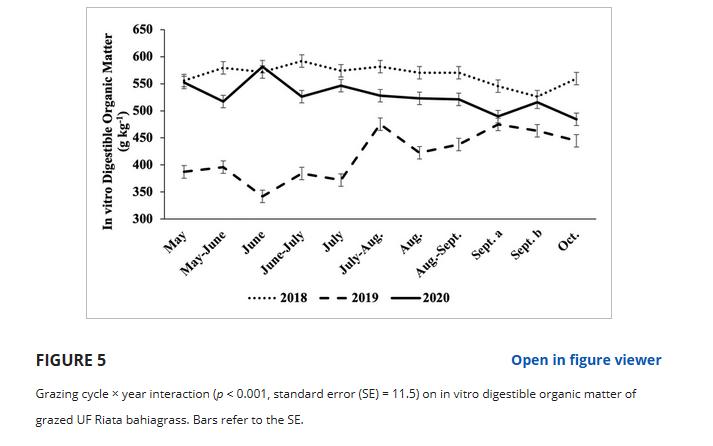
VDOM浓度受处理×年(p = 0.014, SE = 6.8)和放牧周期×年(p < 0.001, SE = 11.5)的影响;图5)交互。
处理×年交互作用的发生是因为HG在2019年的IVDOM浓度高于MG(分别为428和408 g kg - 1), LG是其中的中间(418 g kg - 1)(数据未在图或表中显示)。
然而,在2018年和2020年没有观察到这种差异。
总体而言,2018年的IVDOM (566 g kg - 1)大于2019年(418 g kg - 1), 2020年处于中间水平(526 g kg - 1)。
在所有放牧周期中,IVDOM浓度在年份之间都存在差异,2019年全年的值始终较小(图5)。
5月至7月差异最大,6月差异最大,2019年IVDOM浓度为342 g kg - 1, 2020年为582 g kg - 1。
杂草比例和土壤覆盖率
从HG到LG,随着放牧强度的降低,土壤覆盖度线性增加(p = 0.04),从93%增加到97%(表4)。
UF Riata百喜草草地的总杂草比例在不同处理或年份之间没有变化(p > 0.10);平均而言,杂草在生长季结束时的总贡献为36%。
然而,当单独考虑非种植草和其他杂草的比例时,放牧强度对非种植草的影响呈线性负相关(p = 0.06;表4)。
其他杂草的外观不受处理的影响,但随着时间的推移而增加(p < 0.001;SE = 2)。
2018年,其他杂草的比例为2%,2019年增加到8%,2020年,非草杂草占UF Riata百喜草的22%(数据未显示)。
表4。在放牧季末测定了3种不同放牧强度下处理对百喜草土壤覆盖度、非植草比例和根茎质量的影响。

Root-rhizome生物量
差异有统计学意义(p = 0.09, SE = 1790;表4)和年份(p = 0.07, SE = 2390;数据未在图表或表格中显示)放牧强度对根茎质量的影响。
此外,处理对根茎质量有线性影响(p = 0.03)。
HG的根茎生物量最少,LG的根茎生物量最大,MG介于两者之间。
年份效应的产生是因为根系质量在2019年和2020年(13510 kg hm hm - 1)大于2018年(6130 kg hm hm - 1)。

结论与总结
放牧后残余HM = 800-2200 kg DM ha - 1对UF Riata百喜草总ha和ha率没有影响,这表明该百喜草品种能够耐受广泛的放牧管理。
54.4时CP浓度受影响。2018年、2019年和2020年分别占放牧周期的%、27.3%和36.4%。
当这种差异发生时,HG的CP通常大于LG, MG介于两者之间。
在牧草IVDOM中,HG在某一年优于MG,但IVDOM的差异主要与季节或年份有关。

随着放牧强度的增加,土壤覆被和根茎生物量呈下降趋势,而自发草的比例呈上升趋势。
与LG相比,HG围场的根茎生物量更少,自发草的比例是LG围场的两倍。
相反,非草杂草比例不受放牧强度的影响;然而,这些年来,这一比例从2%上升到了22%。
因此,当UF Riata在作物-牲畜综合系统中用作短期轮作作物时,在2年后终止避免了显著的杂草侵占。
基于我们的研究结果,并考虑到草地持久性和饲料营养价值,我们建议每2周放牧一次的UF Riata百米草的目标放牧后HM为1500 kg DM ha - 1 (MG)。
这样的目标残茬质量可以保持与重度放牧牧场相似的CP浓度值,同时保持与轻度放牧牧场相似的土壤覆盖和根茎生物量。
