前言:
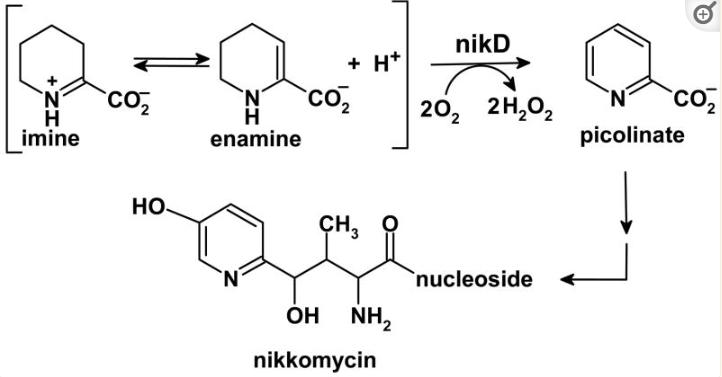
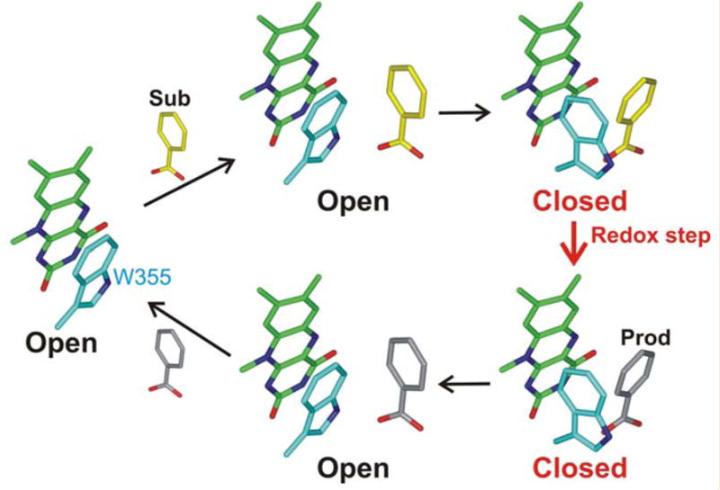
黄素酶nikD是尼霉素抗生素生物合成所必需的,我们发现尼霉素肽的生物合成通过非核糖体途径发生,第一步是由氨基转移酶催化的, 该转移酶将L-赖氨酸转化为α-酮中间体,该中间体环化和脱水以产生哌啶-2-羧酸酯(P2C),一种可以以亚胺和烯胺形式存在的化合物 。
酶纯化和检测
从LB或极好肉汤中培养的细胞(大肠杆菌BL21(DE3)/pDV101)中分离出重组野生型nikD,在LB培养基中培养大肠杆菌BL21(DE3)/pGZ42克隆,该克隆构建了Trp355Phe突变体。
使用与纯化野生型nikD相同的方法分离突变酶,通过测量2纳米处吡啶甲酸盐的形成来测量酶活性,该测定包含264微摩尔P50C,根据黄素含量计算比活性值,并以周转率(μmol吡啶甲酸酯/μmol nikD/min)表示。
通过在2°C下测量不同浓度吡啶甲酸盐的形成,在空气饱和的50毫摩尔磷酸钾缓冲液(pH 8.0)中确定表观稳态动力学参数,纯化的野生型或Trp25Phe突变酶的浓度通过在355纳米处(ε = 455, 11 M^-1 cm^-1)或462纳米处(ε = 12, 700 M^-1 cm^-1)进行吸光度测定确定。
NikD提取物的制备
将NikD在100°C下加热2分钟,以14,000×g的速度离心15分钟,再通过超滤的方式去除变性蛋白质。
通过在66°C下添加*腈乙**(22%,体积/体积)来引起蛋白质的变性,在冰上孵育10分钟后通过离心去除变性蛋白质,进行分析之前,对加热或*腈乙**提取物进行真空浓缩处理。
色谱和光谱
除非另有说明,否则对nikD提取物的HPLC分析在室温下进行,使用配备Hamilton PRP-X100阴离子交换柱(10μm颗粒,4.6mm×250mm)或Microsorb 100-5 C18色谱柱(5μm颗粒、4.6mm×250mm)的Rainin HPLC系统。
柱洗脱液通过在264nm处进行吸光度监测,阴离子交换HPLC分析采用等度洗脱,流速为1.4ml/min或1.2ml/min, 洗脱溶剂由(v/v)1份25mM碳酸铵pH10.3与1份50%甲醇在水(v/v)中混合而成。
使用反相HPLC分析,在先前描述的洗脱方案下,流速为1mL/min,反相HPLC分析采用以下洗脱方案(流速=4mL/min):用溶剂A(5mM乙酸铵,pH10.5)等度洗脱4分钟,进行25分钟的线性梯度至50%溶剂B(90%甲醇/10%水),最后用溶剂A等度洗脱5分钟。
nikD提取物中含有两种ADP衍生物(化合物I和IV),通过使用AMP标准曲线进行定量HPLC分析,确定nikD提取物中化合物I和IV的含量,可用反相柱(化合物I,IV)和/或阴离子交换柱(化合物I)进行分析。
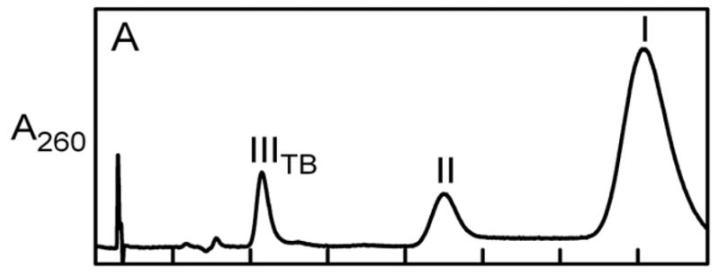
对未处理的配体提取物,根据完整化合物和其AMP分解产物引起的峰下合并面积估算化合物I的含量,在碱性pH下孵育的提取物进行研究时,根据AMP峰下的面积估算化合物I的含量。
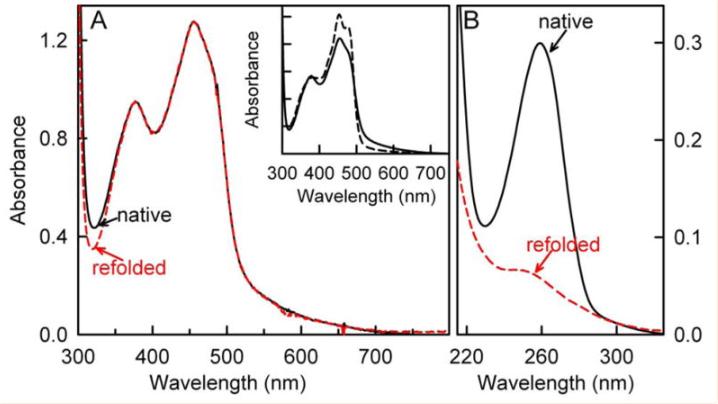
使用Agilent Technologies 8453二极管阵列或PerkinElmer λ 25分光光度计记录吸收光谱,为了确定FAD掺入nikD的化学计量,使用3.0M盐酸胍将酶变性,用专业参数工具计算蛋白质浓度,校正280nm处的吸光度,以排除盐酸胍中游离FAD的贡献(ε280=22,900 M^-1cm^-1)。

质谱分析
NikD配体通过以TB为生长培养基表达的酶的*腈乙**变性释放,LC/MS/MS分析使用微量Q-Tof带有MassLynx工作站的四极杆飞行时间质谱仪进行,配体提取物或标准品经受电喷雾电离(ES)和正/负开关。
除以下所述外,所有HPLC分离均在联盟中进行,使用沃特世亚特兰蒂斯的HPLC系统和dC18色谱柱(3 μm颗粒,2.1 mm×150 mm),进行(流速:250 μL/min),在20 °C下操作。
典型设置为:锥孔电压为20 eV;碰撞能量为1 eV(ESI-MS)或10、15、20(ESI-MS-MS);氩气压力为13磅/平方英寸;离子源温度为125 °C;解离剂温度为290°C。
使用ACQUITY UPLC进行HPLC分离,系统采用PRP-X100阴离子交换柱(5 μm颗粒,4.6 mm×250 mm),在进行单质量数分析以鉴定对应于指定m/z值的可能分子种类时,施加了以下公差限值:公差为11.0 ppm;双键当量(DBE)的最小值为0.0,最大值为25.0;同位素簇参数的分离=1.0,丰度=1.0%。
配体去除NikD的制备
所有步骤均在0至4°C下进行,在10 mL pH 5.0的磷酸钾缓冲液(缓冲液A)中,将含有37 mg nikD的溶液与含有0 mM EDTA和5 M盐酸胍的2.4 mL等分缓冲液A以大约30秒的间隔混合,直到最终盐酸胍浓度达到2 M。
将样品在冰上孵育30分钟后离心,并在含有1 mM EDTA和2 M盐酸胍的缓冲液A中透析过夜,通过透析过程逐渐降低盐酸胍浓度,样品先与含有2 mM EDTA和1.2 M盐酸胍、50.0 M盐酸胍的缓冲液A透析25小时,然后再与缓冲液A透析过夜,对缓冲液A进行2小时透析。
使用Centriprep 1将溶液浓缩至5.30 mL后进行光谱和活性测量,按照上述方法制备复叠蛋白的热提取物,并进行HPLC分析。
Trp355突变为Phe
所有PCR反应均使用Hybaid触地热循环仪进行,PCR产物通过琼脂糖凝胶(1.5%)电泳纯化,用QIAQuick凝胶提取试剂盒回收,测序由MWG Biotech进行。
使用质粒pDV101作为模板,进行1个循环,94°C持续2分钟,左侧片段的正向引物是5′-GACTCAC,后向引物是5′-GAACTTGCC-3′,右侧片段的前向引物是5′-GGGGCGAAGTTC-3′,后向引物是5′-CCGGATATAAGC-3′(END)。

纯化的左手和右手片段分别使用START和END作为正向和反向引物进行合并,(引物中的致突变位点以小写形式显示;密码子355带有下划线),最终PCR产物使用QIAquick PCR纯化试剂盒纯化,并进行NdeI和XhoI酶切,再次纯化。
将其亚克隆到质粒pET23a的NdeI和XhoI位点之间,所得的构建体用于将大肠杆菌BL21(DE3)细胞转化为氨苄青霉素耐药性,为了进行筛选,用QiaPrep旋转迷你制备试剂盒从随机选择的克隆中分离质粒DNA,并用NdeI和XhoI消化。
采用Qiagen质粒Midi试剂盒(Qiagen)分离具有预期插入片段大小(pGZ42)的质粒,并在整个插入片段中进行测序 。
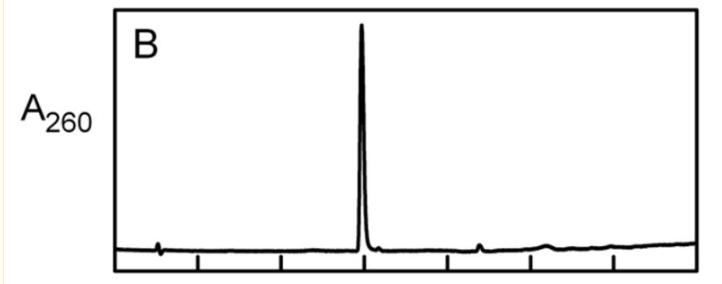
NikD提取物中的内源性配体
我们预计nikD提取物将包含与在晶体结构中的活性位点观察到的内源性配体相对应的单一化合物,通过加热或*腈乙**变性制备的提取物被发现含有至少四种不同的化合物,为了进一步研究这些化合物,使用了阴离子交换或反相柱进行分离。
根据分析,化合物I和IV在结构上相关,并且它们共享一个共同的结合位点,化合物III结合到一个不同的位点,化合物II是化合物I的分解产物。
通过这些结果,可以得出结论,nikD提取物中存在多个化合物,并且它们在结构和结合位点上有所不同。
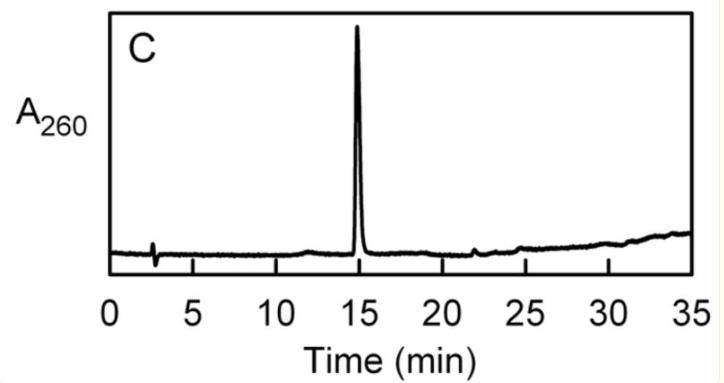
化合物II的鉴定
以下观察结果表明化合物II是AMP,根据我们的观察,化合物II和AMP在HPLC洗脱曲线和吸收光谱方面表现出相似的特征(λ.max= 259 nm)。
此外,当两种化合物经过碱性磷酸酶处理后,它们都被转化为腺苷,并且它们的分子量相同(347 Da),这是通过正或负离子模式下的质谱分析得出的结论。

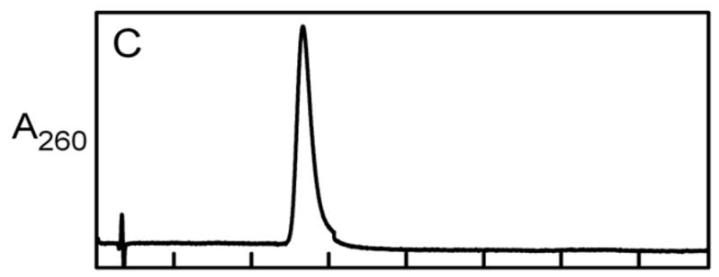
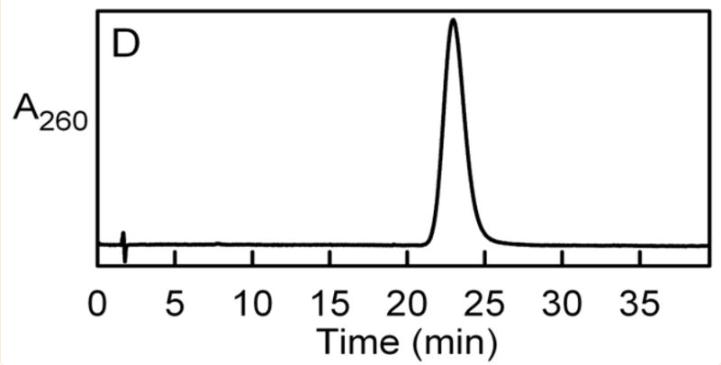
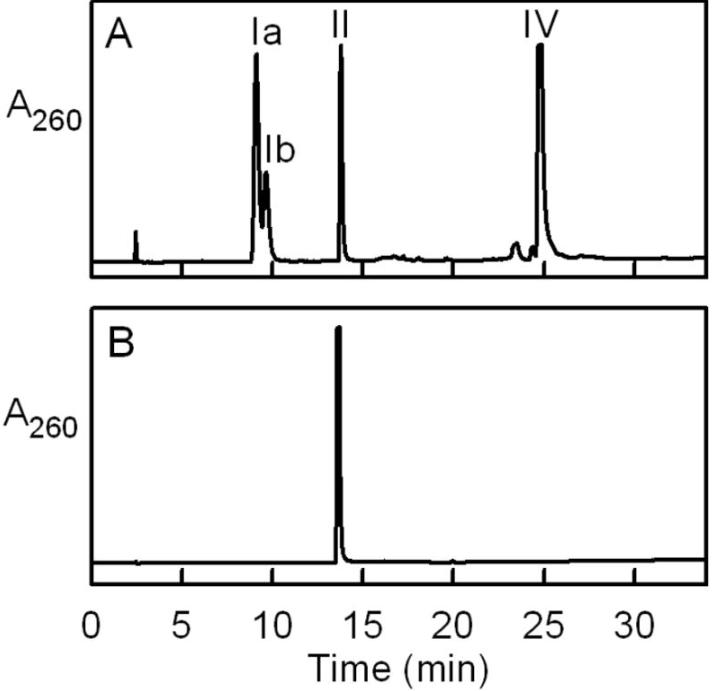
化合物I和IV作为ADP衍生物的性质和鉴定
化合物I是不稳定的,在室温下,1mM KOH中孵育100小时后观察到其定量水解为AMP,化合物在阴离子交换HPLC柱中发生了分离,而在反相洗脱曲线中观察到一对紧密洗脱的峰(Ia,Ib)。
通过反相HPLC的质谱分析发现,化合物Ia和Ib具有相同的分子量,因此它们可能是异构体,与化合物I不同,化合物IV在碱性条件下是稳定的,不容易从阴离子交换柱中回收。
化合物IV很容易通过反相HPLC纯化,纯化的化合物表现出与AMP几乎相同的吸收光谱(λ.max= 259 nm),通过质谱分析确定,化合物IV的分子量是化合物I的两倍以上。
进行正离子模式下的MS/MS分析以鉴定来自化合物I和IV的片段,在使用化合物I和IV进行MS/MS ES谱图时,观察到可归因于AMP、AMP腺嘌呤部分(Ade)以及AMP磷酸化衍生物的信号。
AMP中5'-磷酸盐的磷酸化将产生ADP衍生物,该衍生物应易于被磷酸二酯酶I水解,这与在其他可能位点(例如AMP中的2'-OH或3'-OH)磷酸化产生的化合物不同。
事实上,在使用纯化的磷酸二酯酶I处理粗配体提取物时, 观察到化合物Ia、Ib和IV定量水解为AMP,未检测到其他紫外线吸收产物,这些结果表明,化合物I和IV是ADP衍生物。
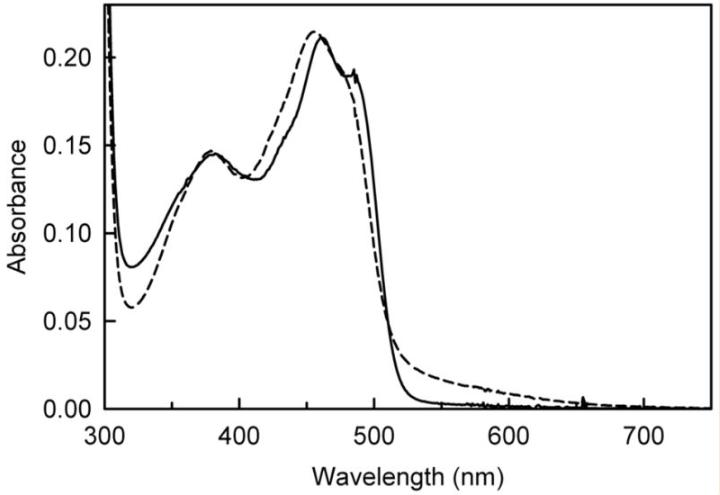
NikD制剂中化合物I和IV的定量分析
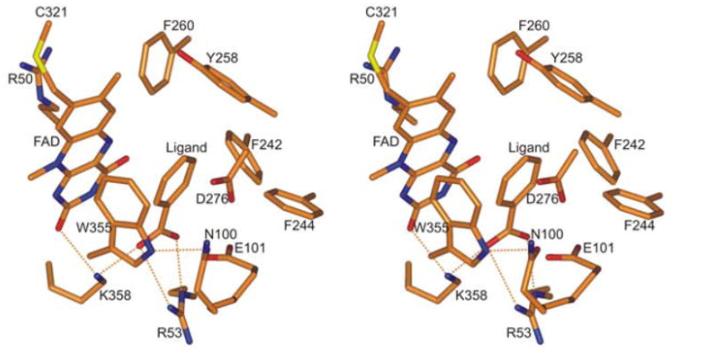
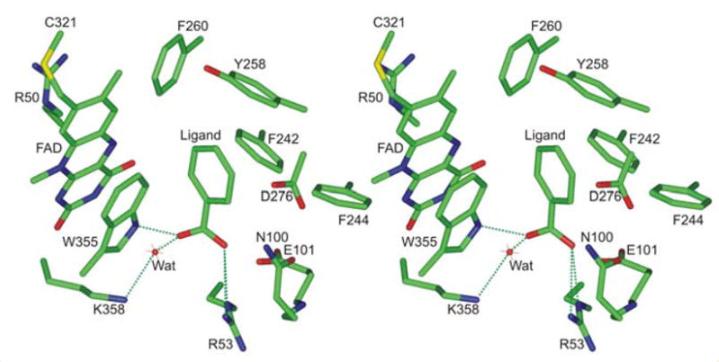
化合物I和IV的大小超过了nikD的活性位点的容纳能力,但可以预期它们可以在黄素结构域的二核苷酸结合基序上结合,并与FAD相互竞争。
在这种情况下,三种化合物的总量不应超过每摩尔nikD一摩尔,为了验证这一假设,使用含有0.54至0.82molFAD的三种酶制剂的提取物进行了定量分析(FAD含量是单独测定的,因为共价结合的黄素不会在酶变性时释放到提取物中)。
化合物I的量在每摩尔nikD的范围为0.02至0.14摩尔之间变化,与FAD的含量无关,相反,化合物IV的量与黄素含量成反比(r2= 0.9589),并且在每摩尔nikD的范围为0.12至0.45摩尔之间变化。
在三种制剂中,FAD、化合物I和化合物IV的摩尔总和在每摩尔nikD中,结果支持了所有三种化合物在nikD中竞争共同ADP结合位点的提议。
Trp355Phe突变体的催化性能
突变体的比活性约为野生型nikD观察到的10%,这是通过使用含有50μM P2C的实验得出的,尽管该底物浓度对于野生型nikD是饱和的,但稳态动力学研究表明,对于表现出明显K的突变蛋白,其饱和度比野生型nikD大30倍。
突变仅导致表观k值降低3倍的半饱和浓度, 这些结果表明,Trp355Phe突变主要影响底物结合,为了评估这一假设,我们通过测量底物类似物与突变酶的结合亲和力来寻找证据。
通过监测酶的可见吸收光谱伴随的扰动,可以轻松测量野生型nikD与各种*制剂抑**复合物的解离常数,对Trp355Phe突变体进行类似研究的多种尝试表明,突变导致酶的稳定性显着降低,这是通过观察在野生型酶长时间稳定的条件下(pH 8.0,25°C或5°C)观察到的聚集现象进行判断的。
作为替代方法,我们比较了1-环己烯酸酯(P1C的2-deaza类似物)对突变型和野生型酶的活性影响,在每种情况下,发现1-环己烯酸酯作为P2C的竞争性*制剂抑**,突变酶的抑制常数与野生型nikD一致,支持了Trp355在底物结合中的假定作用。
结论:
研究表明,在黄酮毒素中,FMN环的外表面和内表面通常分别由酪氨酸和色氨酸残基两侧,色氨酸环相对于黄素环倾斜约20-45°,这种几何形状不利于电荷转移相互作用,酪氨酸残基几乎与黄素环共面,并且当该残基突变为色氨酸(时观察到电荷转移复合物)。
类似地,铁氧还蛋白-NADP还原酶(FNR)中的C端酪氨酸平行于黄素环,将这种酪氨酸更改为色氨酸也会产生一种突变酶,该酶表现出长波长电荷转移带。
参考文献:
【1】乔恩斯,《甲壳素合成酶的微生物*制剂抑**》
【2】 布鲁克纳,《链霉菌Tendae Tu901 L-赖氨酸2-氨基转移酶催化日霉素D生物合成中的初始反应》
【3】卡瑞尔,《nikD的分子表征,nikD是一种在尼霉素抗生素生物合成中很重要的新型黄素酶》
【4】亨特,《在肌氨酸氧化酶中发现第三种辅酶》